


Bosrevue

Het artikel hieronder is een korte samenvatting van de scriptie die de Toekomstboom 2024 won. Deze prijs wordt jaarlijks uitgereikt aan de beste studentenscriptie in het vakgebied van bosecologie en bosbeheer. De prijs is een initiatief van de Stichting Toekomstboom, wier missie het bevorderen is van bosbeheer op wetenschappelijke grondslag, en wordt mogelijk gemaakt door een financiële bijdrage van BOS+, de Stichting Probos, en de Koninklijke Nederlandse Bosbouwvereniging (KNBV). De jury bestond uit vertegenwoordigers van de bosbouwpraktijk uit Nederland en Vlaanderen. De Toekomstboom 2024 werd toegekend aan Anneleen Geussens (KULeuven) voor haar scriptie over de effecten van bosbeheer op droogtestress bij zomereiken en beuken. Overige genomineerden waren Dylan De Greef (KULeuven) met zijn scriptie over de populatiegenetica van Taxus baccata, Stien Melis (KULeuven) met haar scriptie over de rol van mycorrhiza in sterfte door droogte bij grove den en Eline Roelandt (UGent) met haar scriptie “Bosuitbreiding door de blender: verhoogt een mix van spontane en kunstmatige verjonging de klimaatweerbaarheid?”
Onze bossen voorzien ons van hout, nemen CO2 uit de lucht op, brengen ons tot rust… Dit zijn maar een paar voorbeelden van de vele ecosysteemdiensten die bossen ons bieden. Maar in een veranderend klimaat krijgen we steeds vaker te maken met extreme weersomstandigheden, zoals hittegolven en droogtes. Dit is niet alleen onaangenaam voor de mens, maar heeft ook een grote invloed op het functioneren en de vitaliteit van het bos (Dale et al., 2001; Jia et al., 2019). Droge zomers hebben geleid tot meer bladverlies, verminderde groei en hogere sterftecijfers onder bomen in bossen over heel Europa (Sousa-Silva et al., 2018, Taccoen et al., 2019). Tussen 2016 en 2019 hebben maar liefst 100 000 km² Europese bossen schade opgelopen door droogte (European Environment Agency, 2022). Onderzoek naar klimaatslim bosbeheer is noodzakelijk om deze problematiek aan te pakken.

Figuur 1: Zomereiken in Brecht, met symptomen van vitaliteitsverlies als gevolg van droogte in 2018 en 2019 (foto: Sioen et al., 2020).
Dunnen of niet dunnen?
Bossen klimaatslim beheren wordt steeds belangrijker om een bos te creëren dat op een duurzame manier verschillende ecosysteemdiensten kan leveren, en tegelijk veerkrachtiger is, en meer opgewassen tegen een veranderend klimaat met extreme omstandigheden. Een beheerinterventie die mogelijks kan helpen om dit te bereiken, is dunnen. Dit is het selectief vellen van bomen om de dichtheid in het bos te verminderen en zo de groei en gezondheid van de overige bomen te stimuleren (Aussenac, 2000).
Het is echter onduidelijk of dunnen droogtestress in bomen verlicht of net verergert. Sommige onderzoeken tonen aan dat een lagere bosdichtheid door dunning de beschikbaarheid van water in het bosbestand verhoogt doordat er meer water in de bodem kan infiltreren (Giuggiola et al., 2013). Bovendien stimuleert dunnen de groei van het wortelstelsel van de overgebleven bomen, waardoor de boom meer water kan opnemen (Aussenac, 2000). Doordat in een gedund bos minder bomen aanwezig zijn, neemt de competitie voor water eveneens af (D’Amato et al., 2013). Aan de andere kant zorgt een dichter, minder gedund bos voor een kleinere kroon, waardoor de vraag naar water per boom net afneemt (Mausolf et al., 2018). Een hogere stamdichtheid en een meer gesloten kronendak zorgt er eveneens voor dat het bosmicroklimaat beter behouden blijft, waardoor extremen in temperatuur en luchtvochtigheid beter gebufferd kunnen worden (Mausolf et al., 2018). Er bestaat dus bewijs voor zowel de voor- als nadelen van dunnen. De hamvraag in dit onderzoek is dan ook: moeten we onze bossen dunnen om droogteresistentere bossen te creëren? En zo ja, hoe sterk moeten we dan dunnen?
Het onderzoek
Het doel van dit onderzoek is om na te gaan hoe de intensiteit van beheeringrepen zoals dunningen invloed heeft op droogtestress bij bomen. Meer specifiek richtte deze studie zich op zomereiken (Quercus robur) en beuken (Fagus sylvatica) in het Zoniënwoud en het Meerdaalwoud. Hier werd het boomwatertekort (Box 1) opgevolgd in 72 bomen (35 zomereiken en 37 beuken) langsheen een gradiënt van beheerintensiteit gaande van helemaal geen beheer in dichte bosbestanden, tot fel beheer in erg open bosbestanden. De metingen gebeurden tijdens het groeiseizoen van 2022 met behulp van dendrometers (Box 1). Via een statistisch model, dat enerzijds weerfactoren zoals neerslag en luchtvochtigheid, en anderzijds beheerintensiteit combineerde om een stressrespons te voorspellen, kon de beheerintensiteit waarbij minimale stress optreedt, bepaald worden.
Box 1 Boomstammen groeien en krimpen: het boomwatertekort
Droogtestress bij bomen kan worden afgeleid uit dagelijkse fluctuaties in de omtrek van de stam (Zweifel et al., 2005). Er zijn twee zaken met impact op de stamomtrek. Enerzijds zijn er de onomkeerbare veranderingen als resultaat van groei van de boom (Irvine and Grace, 1997). Anderzijds is er ook een omkeerbare component, die een dagelijkse cyclus van inkrimping en uitzetting vormt (Irvine and Grace, 1997). Overdag transpireert de boom via de bladeren water dat opgeslagen zit in de stam, waardoor de stam krimpt. ’s Nachts, wanneer de boom water opneemt uit de bodem, zet de stam weer uit (Vieira et al., 2013). In droogteperiodes kan een boom ‘s nachts niet genoeg water opnemen om die dagelijkse inkrimping te compenseren (Vieira et al., 2013). De hoeveelheid staminkrimping die een boom overdag ondervindt, wordt ook wel het boomwatertekort genoemd, en kan dienen als een maat voor droogtestress (Zweifel et al., 2005).
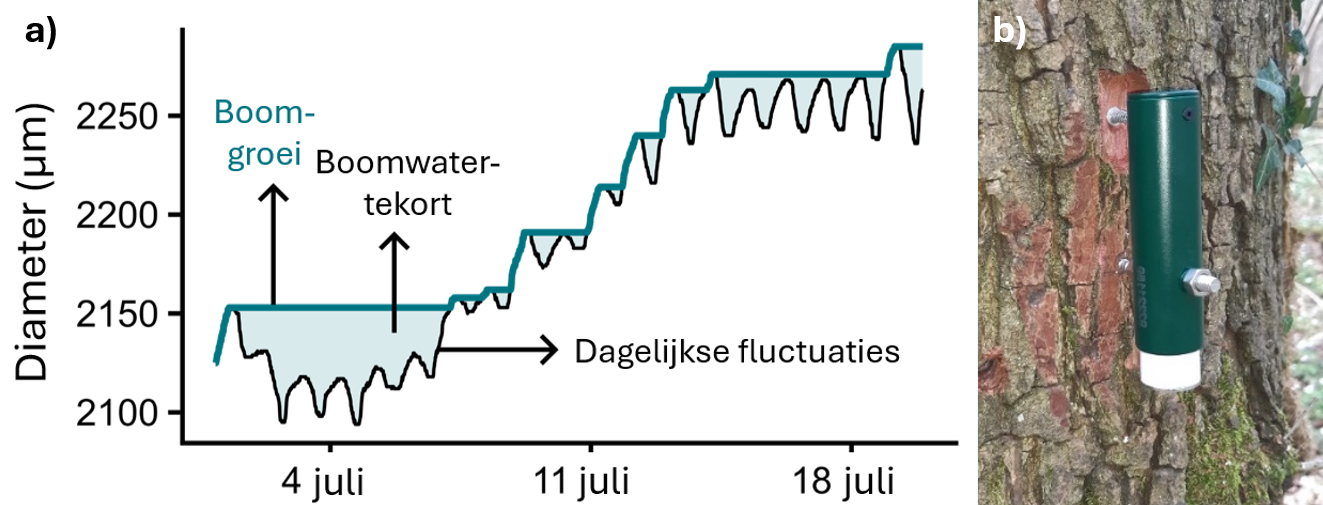
Figuur 2: (a) Typische diameterfluctuaties van een boom. Op lange termijn neemt de diameter toe door groei. Tegelijk kent de boomstam een dagelijkse cyclus van inkrimping en uitzetting, doordat de boom overdag transpireert, en ’s nachts weer water uit de bodem opneemt. (b) Een dendrometer van het gebruikte type, bevestigd aan een zomereik (Foto: Anneleen Geussens).
Deze dagelijkse fluctuaties kunnen worden gemeten met een dendrometer. Dit apparaat wordt aan een boom bevestigd, waarna het met intervallen van 15 minuten tot op een micrometer nauwkeurig de stamfluctuaties meet.
Verschillende soorten, verschillende voorkeuren
Dit onderzoek toonde enerzijds dat in de lente van 2022, waar de droogteperiode lang aanhield maar niet erg intens was door de hoge luchtvochtigheid, zomereik een sterkere stressrespons dan beuk vertoonde. Anderzijds vertoonde beuk meer stress dan zomereik in de zomer van 2022, die gekenmerkt was door een aantal korte en intense droogteperiodes. Dit bevestigt dat eiken gevoelig zijn aan langere droogtes, terwijl beuken meer lijden onder intense droogte met lage luchtvochtigheid. Dit heeft te maken met de waterstrategie van deze twee soorten. In een droge periode sluiten eiken de huidmondjes in hun bladeren om overmatige verdamping van water tegen te gaan (Tardieu and Simonneau, 1998; Thomsen et al., 2019). Hierdoor kan echter ook geen CO2 meer opgenomen worden, waardoor de boom ‘verhongert’ door een koolstoftekort op langere termijn: zonder opname van CO2 kan immers geen fotosynthese plaatsvinden, en dus ook geen opbouw van organische stoffen en geen aanleg van reserves (McDowell et al., 2008; Thomsen et al., 2019). Beuken daarentegen, zullen hun huidmondjes open houden, om zulke ‘koolstofverhongering’ te voorkomen, maar verliezen hierdoor tijdens droogtes veel water (Tardieu and Simonneau, 1998; McDowell et al., 2008). Bovendien heeft eik, als lichtboomsoort, een hogere droogtetolerantie dan de schaduwtolerante beuk (Niinemets en Valladares, 2006).
Dit verschil in waterstrategie tussen de twee soorten, zorgt ook voor een verschillende respons op beheerintensiteit. Droogtestress bij zomereiken neemt af naarmate de beheerintensiteit toeneemt. Bij een sterker beheer zijn er immers minder bomen in het bos aanwezig, waardoor concurrentie voor water tussen de bomen afneemt (Giuggiola et al., 2013; D’Amato et al., 2013). Wordt de beheerintensiteit echter té hoog, dan neemt de droogtestress weer toe. Eiken ondervonden het minste droogtestress in bosbestanden met een totale oppervlakte van stamdoorsneden tussen 13 en 21 m²/ha.
Voor beuk, daarentegen, leidt een toename in beheerintensiteit steeds tot een toename in droogtestress. Hoewel sterk beheer de concurrentie in het bos sterk vermindert, krijgt het bos ook een opener karakter, waardoor het microklimaat in het bos verzwakt (Mausolf et al., 2018). Dit betekent dat het klimaat in het bos meer gevoelig is aan schommelingen en extremen in temperatuur en luchtvochtigheid buiten het bos. Een bos met een minder uitgesproken microklimaat zal periodes van extreme droogte en hitte dus feller voelen dan een bos met een sterk uitgesproken microklimaat. Beuk is bijzonder gevoelig voor deze extremen. Om droogtestress tot een minimum te beperken heeft beuk dus een voorzichtig beheer nodig. Beuken ondervonden het minste droogtestress in bestanden met een grondvlak tussen 24 en 33 m²/ha. Ter vergelijking, de onbeheerde beukenbestanden die deel uitmaakten van het onderzoek hadden een grondvlak tussen 28 en 33 m²/ha.

Figuur 3: Schematische voorstelling van de effecten van beheer in het bos. Een toenemende beheerintensiteit zorgt voor een afname van concurrentie en een toename van beschikbaarheid van water. Bij intenser beheer neemt echter ook de buffering van het microklimaat af, waardoor het bos gevoeliger wordt voor extreme droogte en hitte.
Beheer tegen droogtestress
De contrasterende respons van de twee soorten op beheerintensiteit, toont dat verschillende beheerstrategieën aangewezen zijn om droogteresistentie te optimaliseren: een sterker beheer met lagere stamdichtheid voor zomereik, en een lichter beheer met hogere stamdichtheid voor beuk. Een casestudy in het Meerdaalwoud toont echter dat de situatie op terrein verschilt van deze optima. De meeste zomereikenbestanden in het Meerdaalwoud zijn te dicht om de droogtestress te minimaliseren (Agentschap voor Natuur en Bos, 2004). Van de beukenbestanden ligt ongeveer de helft binnen de optimale bestanddichtheid voor minimale stress, terwijl de andere helft te open is (Agentschap voor Natuur en Bos, 2004). Een meer gedifferentieerde beheerstrategie voor deze twee soorten kan dus helpen om droogtestress te minimaliseren in het Meerdaalwoud.
Bij de keuze van de beheerintensiteit, mogen andere aspecten dan droogte-resistentie, zoals houtkwaliteit, biodiversiteit en stabiliteit, echter niet vergeten worden. Het is cruciaal om een juiste balans te vinden tussen al deze aspecten die door het bosbeheer beïnvloed worden. Bovendien zijn de resultaten van dit onderzoek gebaseerd op maar één droogtejaar, in monocultuurbossen. Om een concreet en consistent beheeradvies te kunnen geven, is het nodig om de bomen over meerdere jaren en meerdere droogteperiodes op te volgen, alsook de invloed van diversiteit in het bos te bestuderen.
Dit onderzoek biedt inzichten in hoe een goede keuze van beheerintensiteit kan bijdragen aan het minimaliseren van droogtestress bij bomen. Dit is een dringende zaak in een veranderend klimaat, waarin extreme droogtes steeds vaker voorkomen en bossen een steeds belangrijkere rol spelen bij het mitigeren van klimaatverandering en de gevolgen ervan. In deze kritieke omstandigheden kan het optimaliseren van de beheerintensiteit dienen als middel om de droogtestress in het veranderende klimaat te bestrijden.
Tot slot
Tot slot willen we de geïnteresseerde lezer voor meer details graag verwijzen naar de scriptie die de basis vormt voor dit artikel (Geussens A. (2023). The effects of forest management intensity on drought stress in pedunculate oak and European beech. [Masterproef, KU Leuven]. KU Leuven e-thesissen.), alsook het artikel door Verdonck et al. (2025), dat op deze bevindingen voortbouwt (Verdonck S., Geussens A, Zweifel R., Thomaes A., Van Meerbeek K., Muys B. (2025). Mitigating drought stress in European beech and pedunculate oak: The role of competition reduction. Forest Ecosystems 13.)
Gelieve als volgt te citeren:
Anneleen Geussens (2025) Dunnen of niet dunnen? De effecten van bosbeheer op droogtestress bij zomereiken en beuken. Bosrevue 121a.
ISSN 2565-6953 – Bosrevue 121a
-
Referenties
Agentschap voor Natuur en Bos. (2004). Bestandsfiches Heverleebos en Meerdaalwoud. [online]. Beschikbaar via: https://www.natuurenbos.be/sites/default/files/bijlage_bestandsfiches_ heverleebos_meerdaalwoud.pdf. [Geraadpleegd op 11/04/2023].
Aussenac G. (2000). Interactions between forest stands and microclimate: Ecophysiological aspects and consequences for silviculture. Annals of Forest Science 57: 287-301.
Dale V.H., Joyce L.A., McNulty S., Neilson R.P., Ayres M.P., Flannigan M.D., Hanson P.J., Irland L.C., Lugo A.E., Peterson C.J., Simberloff D., Swanson F.J., Stocks B.J., Wotton B.M. (2001). Climate Change and Forest Disturbances: Climate change can affect forests by altering frequency, intensity, duration, and timing of fire, drought, introduced species, insect and pathogen outbreaks, hurricanes, windstorms, ice storms, or landslides. BioScience 51(9): 723-734.
D’Amato A.W., Bradford J.B., Fraver S., Palik B.J. (2013). Effects of thinning on drought vulnerability and climate response in north temperate forest ecosystems. Ecological Applications 23(8): 1735-1742.
European Environment Agency. (2022). Drought impact on ecosystems in Europe. [online]. Beschikbaar via: https://www.eea.europa.eu/ims/drought-impact-on-ecosystems-in-europe. [Geraadpleegd op 08/04/2022].
Giuggiola A., Bugmann H., Zing A., Dobbertin M., Rigling A. (2013). Reduction of stand density increases drought resistance in xeric Scots pine forests. Forest Ecology and Management 310: 827-835.
Irvine J., Grace J. (1997). Continuous measurements of water tensions in the xylem of trees based on the elastic properties of wood. Planta 202: 455-461.
Jia G., Shevliakova E., Artaxo P., De Noblet-Ducoudré N., Houghton R., House J., Kitajima K., Lennard C., Popp A., Sirin A., Sukumar R., Verchot L. (2019). Land–climate interactions. In: Climate Change and Land: an IPCC special report on climate change, desertification, land degradation, sustainable land management, food security, and greenhouse gas fluxes in terrestrial ecosystems [Shukla P.R., Skea J., Calvo Buendia E., Masson-Delmotte V., Pörtner H.-O., Roberts D.C., Zhai P., Slade R., Connors S., van Diemen R., Ferrat M., Haughey E., Luz S., Neogi S., Pathak M., Petzold J., Portugal Pereira J., Vyas P., Huntley E., Kissick K., Belkacemi M., Malley J., (eds.)]. Intergovernmental Panel on Climate Change.
Mausolf K., Wilm P., Härdtle W., Jansen K., Schuldt B., Sturm K., von Obheimb G., Hertel D., Leuschner C., Fichtner A. (2018). Higher drought sensitivity of radial growth of European beech in managed than in unmanaged forests. Science of the Total Environment 642: 1201-1208.
McDowell N., Pockman W.T., Allen C.D., Breshears D.D., Cobb N., Kolb T., Plaut J., Sperry J., West A., Williams D.G., Yepez E.A. (2008). Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. New Phytologist 178(4): 719-739.
Niinemets Ü., Valladares F. (2006). Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecological Monographs 76(4): 521-547.
Sioen G., Verschelde P., Roskams P. (2020). Bosvitaliteitsinventaris 2019. Resultaten uit het bosvitaliteitsmeetnet. Rapporten van het Instituut voor Natuur- en Bosonderzoek 2020 (20). Instituut voor Natuur- en Bosonderzoek, Brussel.
Sousa-Silva R., Verheyen K., Ponette Q., Bay E., Sioen G., Titeux H., Van de Peer T., Van Meerbeek K., Muys B. (2018). Tree diversity mitigates defoliation after a drought-induced tipping point. Global Change Biology 24: 4304-4315.
Taccoen A., Piedallu C., Seynave I., Perez V., Gégout-Petit A., Nageleisen L., Bontemps J., Gégout J. (2019). Background mortality drivers of European tree species: climate change matters. Proceedings of the Royal Society B: Biological Sciences 286(1900).
Tardieu F., Simonneau T. (1998). Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviour. Journal of Experimental Botany 49: 419-432.
Thomsen S., Reisdorff C., Gröngröft A., Jensen K., Eschenbach A. (2019). Responsiveness of mature oak trees (Quercus robur L.) to soil water dynamics and meteorological constraints in urban environments. Urban Ecosystems 23: 173-186.
Vieira J., Rossi S., Campelo F., Freitas H., Nabais C. (2013). Seasonal and daily cycles of stem radial variation of Pinus pinaster in a drought-prone environment. Agricultural and Forest Meteorology 180: 173-181.
Zweifel R., Zimmerman L., Newbery D.M. (2005). Modelling tree water deficit from microclimate: an approach to quantifying drought stress. Tree Physiology 25: 147-156.


