


Bosrevue

Het artikel hieronder is een korte samenvatting van de scriptie die de Toekomstboom 2023 won. Deze prijs wordt jaarlijks uitgereikt aan de beste studentenscriptie in het vakgebied van bosecologie en bosbeheer. De prijs is een initiatief van de Stichting Toekomstboom, wier missie het bevorderen is van bosbeheer op wetenschappelijke grondslag, en wordt mogelijk gemaakt door een financiële bijdrage van BOS+, de Stichting Probos, en de Koninklijke Nederlandse Bosbouwvereniging (KNBV). De jury bestond uit vertegenwoordigers van de bosbouwpraktijk uit Nederland en Vlaanderen. De Toekomstboom 2023 werd toegekend aan Louis Verschuren (Universiteit Gent) voor zijn scriptie over de reactie van jonge en oude beuken op klimaatverandering. Overige genomineerden waren Dirk Hemmelder (Van Hall Larenstein) met zijn scriptie “CO2 certification of new forests” en Marte Vanlangenaeker (KU Leuven) met haar scriptie “Artificiële bosverjonging via kloempen in de Vlaamse en Nederlandse zandstreek”.
Beuk (Fagus sylvatica) is een van de meest voorkomende loofboomsoorten in de Europese bossen en is van nature de dominante boomsoort in het grootste deel van België. Een honderdtal beukenbossen in 18 Europese landen (waaronder een deel van het Zoniënwoud) zijn beschermd als UNESCO-werelderfgoed omwille van hun unieke natuurwaarde en kolonisatiegeschiedenis in Europa sinds de laatste ijstijd.
Volgens sommige wetenschappers dreigt klimaatverandering echter de dominante positie van beuk onder druk te zetten (Allen et al., 2015; Reyer et al., 2013). Vele onderzoekers benadrukken bijvoorbeeld de droogtegevoeligheid van beuk (Brinkmann et al., 2019; Cavin & Jump, 2017; Köcher et al., 2009; Leuschner, 2020; Leuschner and Wedde, 2019; Scherrer et al., 2011). Andere bronnen benadrukken echter de veerkracht van beuk, of relativeren deze impact, en maken die sterk afhankelijk van het gevolgde klimaatscenario (Dyderski et al., 2018; Leuschner, 2020; Martinez del Castillo et al., 2022).
De klimaatmodellen voor België voorspellen naast hogere gemiddelde temperaturen ook een minder uniforme verdeling van regen met nattere winters en drogere zomers tot gevolg (Campioli et al., 2012; Termonia et al., 2018). Extreme klimatologische gebeurtenissen zoals droogtes en hittegolven zullen ook vaker voorkomen en intenser zijn. Deze voorspelde omstandigheden kunnen grote gevolgen hebben voor de vitaliteit en groei van beuk aangezien deze gevoelig is aan langdurige droogte, vooral in de lente en de zomer (Giagli et al., 2016; van der Werf et al., 2007). Dit vooral omdat de beuk een oppervlakkig wortelstelsel en groot bladoppervlak heeft, wat nadelig is tijdens periodes van lange droogte. Een oppervlakkig wortelstelsel kan minder putten uit diepere grondwatervoorraden. Een groot bladoppervlak betekend meer waterverdamping (transpiratie) en dus een hoger verbruik van water.
De grootste beuken ter wereld zijn in het Zoniënwoud te vinden (Vandekerkhove et al., 2018). De groei van de beuken op deze UNESCO-werelderfgoed site zijn het voorwerp van dit onderzoek. Hierbij wordt zowel gekeken naar levende bomen, als naar individuen die recent omgewaaid (‘windgeworpen’) zijn. Zo kunnen we een beeld vormen van de verschillen tussen beide groepen in hun groei en hun reactie op een veranderend klimaat. Aangezien beuk vaak oppervlakkig wortelt is het omwaaien van deze boom – een zogenaamde windworp – de belangrijkste doodsoorzaak bij oudere individuen. De hypothese is dat windgeworpen beuken een ander groeipatronen vertonen in de jaren voor hun val. We onderzoeken ook wat de impact van zaadproductie is op de groei (vaker voorkomen van mastjaren, jaren met exceptioneel hoge vruchtproductie).
Materialen en methoden
In dit onderzoek wordt de groei van oude beuken in het Zoniënwoud heel gedetailleerd in beeld gebracht aan de hand van jaarringanalyses. De onderzochte bomen bevinden zich in het onderzoeksbestand van het bosreservaat Joseph Zwaenepoel (zie coverfoto). Dit bestand is ongeveer 15 ha groot, werd aangeplant rond 1775, en is al sinds 1983 uit beheer genomen (Baeté et al., 2002). Het bestand valt op door zijn hoge dichtheid aan zeer grote beuken en grote volumes dood hout. De recordboom in dit bestand heeft zelfs een diameter van 167 cm en een volume van 59 m³. Deze boom is daarmee één van de grootste levende beuken (in volume) ter wereld (Demol et al., 2020).
In het kader van dit onderzoek werden 58 oude beuken aangeboord: 39 nog levende bomen en 16 recent omgewaaide bomen (zie Figuur 1). Uit de stam van de geselecteerde bomen werden telkens twee potloodvormige staafjes gehaald, met behulp van een lange aanwasboor (Presslerboor, zie Haneca (2017)). Op deze boorspanen zijn alle jaarringen van de bast tot in de kern te zien. Deze staalname is een zeer kleine ingreep en heeft geen impact op de gezondheid van de levende bomen (Haneca, 2017; Speer, 2010; Tsen et al., 2016). Aan de hand van hun jaarringen kan er onderzocht worden hoe deze bomen reageren op extreme klimatologische gebeurtenissen (zoals droogtes) en of er een verschil in groei te vinden is tussen nog levende bomen en recent windgeworpen bomen in de jaren voor hun val.

Figuur 1: De twee bemonsterde groepen van beuken die tot 250 jaar oud zijn: nog levende bomen (links) en recent omgewaaide bomen (rechts). Op de foto’s is ook de bemonstering met een presslerboor te zien.
De gemaakte boorstalen werden gescand aan de hand van X-stralen microtomografie (De Mil et al., 2016; Van den Bulcke et al., 2019; Van den Bulcke et al., 2014). Deze techniek laat toe om in hoge resolutie digitale 3D beelden te maken waarop de dichtheid van het hout en dus de jaarringen goed te zien zijn. Op die manier kunnen we “in” de boorkern kijken zonder deze te moeten versnijden. Op figuur 2 zijn er 2D slices uit deze 3D beelden te zien.
Vervolgens werden op de scans de jaarringgrenzen aangeduid. Een voorbeeld van dergelijke jaarringen is te zien in Figuur 2. In totaal werden voor dit onderzoek op de 116 gemaakte boorkernen meer dan 20.000 jaarringen opgemeten. Met deze jaarringdata kunnen dan zowel de groeitrends als de droogtegevoeligheid van deze bomen onderzocht worden.
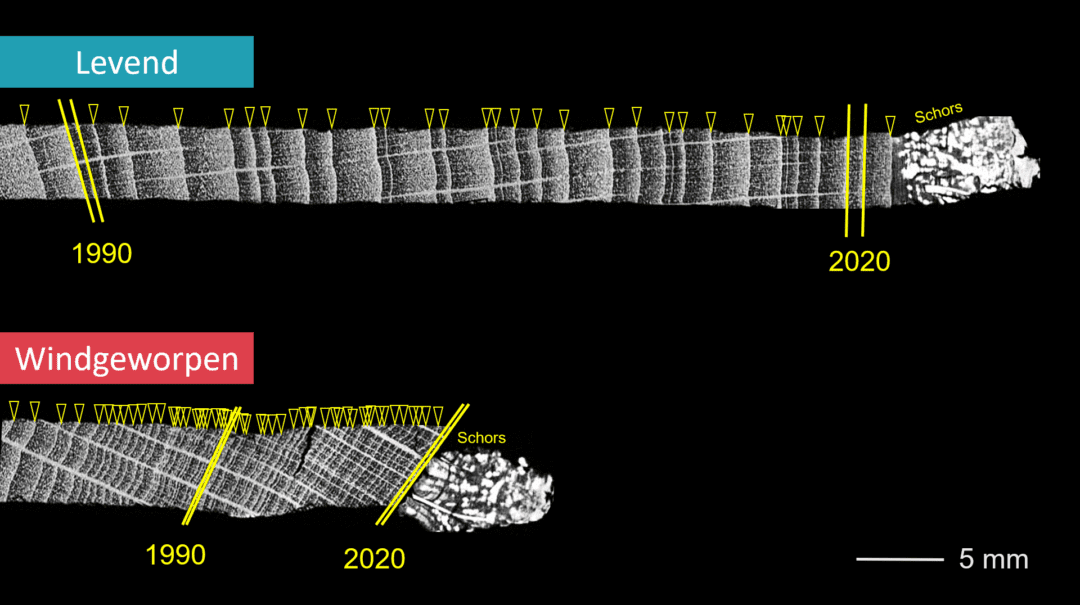
Figuur 2: Een voorbeeld van twee langwerpige boorstalen uit dit onderzoek met beeldvorming aan de hand van XµCT. Dit zijn 2D slices uit de gemaakte 3D beelden. De grijswaarden geven de densiteit weer van licht hout in het voorjaar (zwart) tot dens hout in het najaar (wit). De jaarringgrenzen op deze figuur zijn aangeduid met gele driehoekjes bovenaan elk staal. Er is een groot recent groeiverschil te zien tussen deze twee bomen (de schaal is dezelfde in beide foto’s). In het bovenste staal zijn duidelijke afwisselingen van veel en weinig groei (grote en kleine ringen) te zien. In het onderste staal zijn de jaarringen overwegend smal, er was dus een trage groei.
Groei doorheen de jaren
De jaarlijkse groei wordt het best beschreven via de jaarlijkse aanwas van het grondvlak (de zogenaamde basal area increment, BAI). Dat is de jaarlijkse toename in oppervlakte van het grondvlak (stamoppervlak van de boom op 1,3 m hoogte). Deze maat houdt immers niet alleen rekening met de breedte van de groeiring, maar ook met de grootte van de boom. Het is een maat voor de groei waarbij de steeds toenemende diameter van de boom in rekening gebracht wordt (Goris, 2014). Een groeiring van 1 cm bij een jonge (en dus nog slanke) boom vertegenwoordigt immers veel minder biomassa dan eenzelfde groeiring van 1 cm bij een oude dikke boom. Het verloop van de BAI per boom is te zien in Figuur 3.
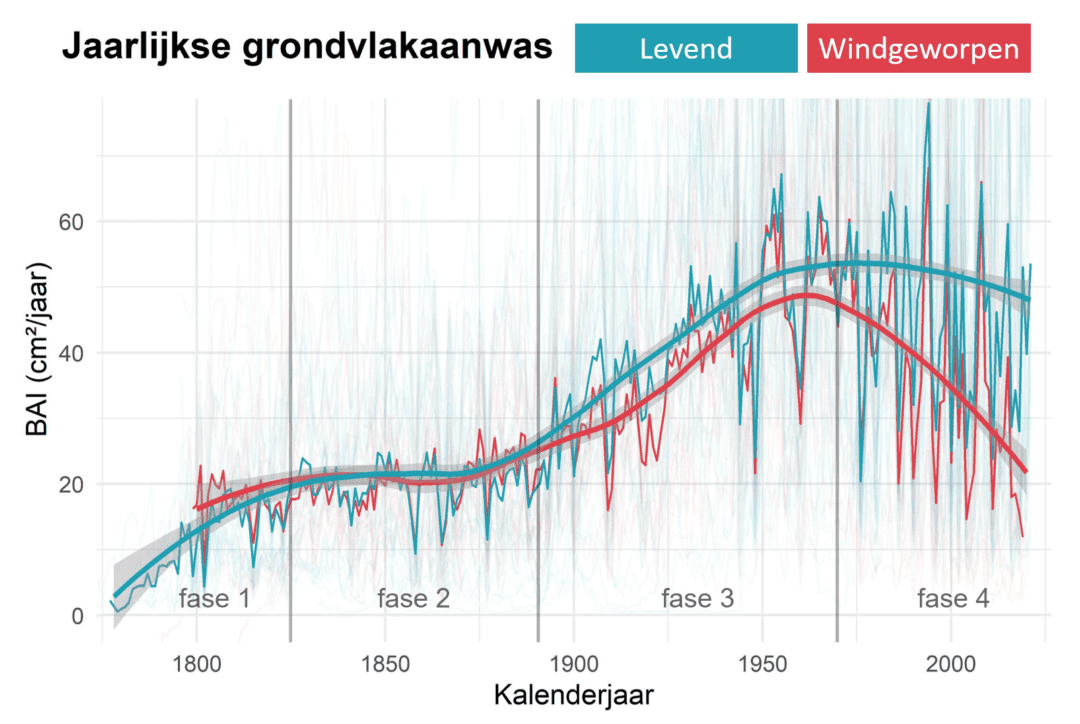
Figuur 3: De jaarlijkse grondvlakaanwas (BAI) van de twee onderzochte groepen doorheen de tijd. Zowel de ruwe data (in het bleek), het gemiddelde per jaar (volle rode en blauwe lijn), en de algemene trend (dikke rode en blauwe lijn) zijn geplot. (Figuur aangepast uit wetenschappelijk artikel, zie link onderaan)
De groei van deze beuken kan opgedeeld worden in 4 fases:
- Fase 1 (1775 – 1825): de initiële groei met een stijgende trend in BAI van 0 naar 20 cm²/jaar
- Fase 2 (1825 – 1890): een afvlakking van de groei met vrij constante BAI rond 20 cm²/jaar
- Fase 3 (1890 – 1970): een sterke stijging van de groei waarbij de BAI steeg van 20 naar gemiddeld 55 cm²/jaar. Meer dan een verdubbeling van de groei dus.
- Fase 4 (1970 – 2021): bij de nog levende bomen een afvlakking tot lichte daling van 57 naar 52 cm²/jaar. Bij de recent windgeworpen bomen is dit echter een sterke groeidaling van 50 naar minder dan 25 cm²/jaar. Wat opvallend is: de afwijkende trend ten opzichte van de levende bomen zet zich al in vanaf de jaren 1970. Deze recent omgevallen beuken hadden het dus al ruim 40 jaar opvallend moeilijker dan de nu nog levende bomen die in diezelfde periode maar weinig in groeisnelheid gedaald zijn. Deze laatste fase wordt in beide groepen ook getypeerd door zeer grote schommelingen in de groei waarbij jaren met zeer brede jaarringen worden opgevolgd door jaren met zeer weinig groei (en dus heel smalle jaarringen).
Groei in tijden van droogte
Zoals eerder aangegeven kan droogte tijdens het groeiseizoen de groei en aanmaak van nieuw hout sterk beïnvloeden. De mate waarin de bomen hiervan herstellen in de daaropvolgende jaren is dan weer een aanwijzing voor hun herstelvermogen (‘resilience’). Om dit te kwantificeren hebben we de groei tijdens de 10 droogste voorjaren (maart tot mei) en 10 droogste zomers (juni tot augustus) van afgelopen 50 jaar, en van de daaropvolgende twee jaren, uitgezet tegenover de gemiddelde groei rond die periode (zogenaamde Ring Width Index, RWI), zowel voor de nog levende als de windgeworpen bomen. Dit is weergegeven in Figuur 4. Een RWI van 1 (rode stippellijn) betekent dat de groei op een gemiddeld niveau zat tijdens dat jaar. Hoe lager de RWI, hoe sterker de groei negatief afweek van de gemiddelde groeitrend.
Een droogte tijdens het voorjaar had bij de nog levende bomen vooral het jaar zelf een negatieve impact: ze groeiden gemiddeld 15% minder dan de jaren ervoor. Het jaar na de droogte was de groei echter alweer hersteld tot op het niveau van voor de droogte. Een droogte tijdens de zomer had het jaar zelf geen impact op de groei van de nog levende bomen; deze bomen groeiden even goed als in de jaren ervoor. Het jaar na die zomerdroogte was er echter wel een negatief effect: de levende bomen groeiden dan 10% minder dan in de jaren voor de droogte. Belangrijk hierbij is dat deze bomen dus weldegelijk gevoelig zijn aan droogtes, maar dat ze opvallend snel herstellen na zo een moeilijk jaar. Deze bomen vertonen dus een opvallende veerkracht. Ook opmerkelijk: windgeworpen bomen reageerden niet significant slechter op extreme droogtes. Hoewel er dus een verschil was in algemene groeitrend, vinden we geen verschil in droogtegevoeligheid tussen levende en windgeworpen beuken.
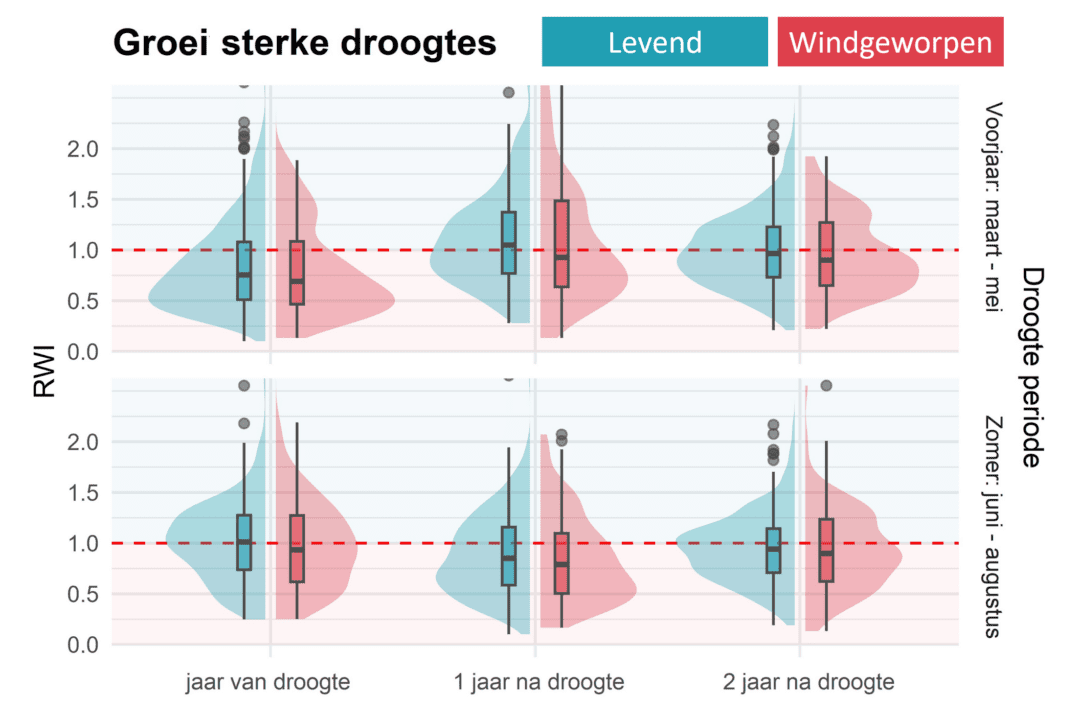
Figuur 4: (foto4.PNG) De groei tijdens en na een droogte in het voorjaar (boven) en in de zomer (onder). Het gaat telkens om de 10 droogste jaren van de laatste 50 jaar. De groei is hier uitgedrukt als RWI (Ring Width Index). Een RWI van 1 (rode stippellijn) betekent dat de groei op een gemiddeld niveau zat tijdens dat jaar. Hoe lager de RWI, hoe sterker de groei negatief afweek van de algemene groeitrend. (Figuur aangepast uit wetenschappelijk artikel, zie link onderaan)
Veel vruchten, weinig hout
Hoge zomertemperaturen en veel zonnestraling kunnen bij beuk zorgen voor een mastjaar (jaar met zeer hoge productie van vruchten) in het daaropvolgende jaar (Hacket-Pain et al., 2015; Leuschner, 2020; Müller-Haubold, 2015). Tijdens een mastjaar kan één derde tot de helft van de suiker aangemaakt in de fotosynthese gebruikt worden voor het produceren van vruchten waardoor er minder energie over is om bij te dragen aan groei (Dittmar et al., 2003; Drobyshev et al., 2010; Müller-Haubold et al., 2013). Mastjaren komen steeds vaker voor door de toegenomen frequentie van hittegolven en aanhoudende atmosferische depositie van stikstof
(Drobyshev et al., 2010; Latte et al., 2015; Müller-Haubold et al., 2015; Vacchiano et al., 2017; Vanhellemont et al., 2019).
De jaarlijks gevormde hoeveelheid beukenzaad wordt in het Zoniënwoud gemeten sinds 1999. Dit wordt gedaan binnen het ICP Forests Level II proefvlak, dat vlakbij de onderzoeksplot gelegen is. Deze zaadproductie wordt in figuur 5 weergegeven (als bruine balken, op de linker Y-as), samen met de jaarlijkse groei (RWI) van de onderzochte bomen (als gekleurde lijnen, op de rechter Y-as). Er is bij beide groepen een duidelijke negatieve correlatie tussen groei en zaadproductie: zo was er tijdens mastjaren 35% minder groei dan in niet-mastjaren. Ook hier reageren windgeworpen bomen niet significant slechter op mastjaren dan nog levende bomen.
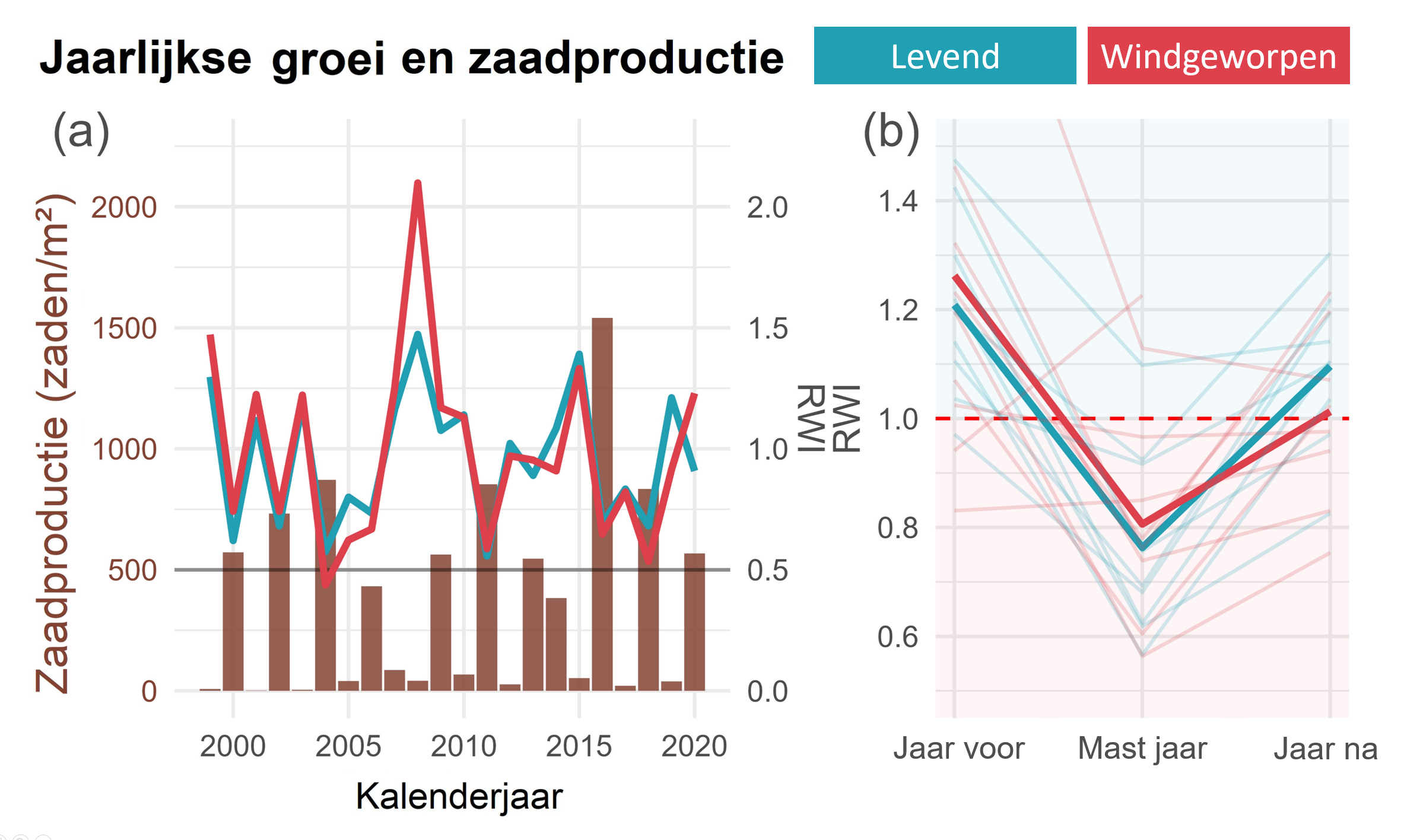
Figuur 5: (foto5.PNG) (a) De jaarlijkse groei (RWI) van de twee populaties doorheen de tijd (gekleurde lijnen) met eronder de jaarlijkse zaadproductie (bruine balken). De grens van wat wel of niet een mastjaar is staat op 500 zaden/m² (zwarte horizontale lijn). We zien dat bij mastjaren de groei sterk afneemt. (b) de gemiddelde groei (RWI) voor, tijdens, en na elk mastjaar tussen 1999 en 2020. De aparte jaren zijn aangegeven als lichte lijnen. (Figuur aangepast uit wetenschappelijk artikel, zie link onderaan)
Discussie
De sterke stijging van de groei tussen 1890 en 1970 waarbij de BAI meer dan verdubbelde (zie Figuur 3) kan naast beheer en leeftijdseffecten ook toegeschreven worden aan de stijging van stikstofdepositie in die periode. Bossen werden nutriënten-rijker en groeiden hierdoor sterker (Laubhann et al., 2009). De sterke schommeling van de groei vanaf 1970 (zie Figuur 3) is waarschijnlijk voornamelijk te wijten aan het voorkomen van meer en intensere droogteperiodes die elkaar sneller opvolgen in combinatie met vaker voorkomende mastjaren (Drobyshev et al., 2010; Latte, 2017; Latte & Claessens, 2015; Leuschner, 2020; Vacchiano et al., 2017; Vanhellemont et al., 2019).
De recent omgewaaide bomen hadden, in tegenstelling tot de nog levende bomen, een sterk dalende groeitrend sinds de jaren ‘70 (zie Figuur 3). Het verschil tussen de levende en windgeworpen bomen manifesteert zich dus al heel lang, waarbij we kunnen stellen dat deze bomen al 30-40 jaar ‘aan het vallen’ zijn. Er werden echter geen significante verschillen in droogtegevoeligheid gevonden. De dalende groei bij omgewaaide beuken valt mogelijks te verklaren door verschillen in vitaliteit van het wortelstelsel. Door de steeds vaker voorkomende droogtes moeten deze bomen veel energie besteden aan het herstel van bladeren en kleine takken. Als daarbovenop om de twee jaar een grote hoeveelheid zaad geproduceerd moet worden, blijft er maar weinig energie over om, bijvoorbeeld, beschadigde wortels te herstellen. We veronderstellen dat de windgeworpen bomen een minder ontwikkeld wortelstelsel hadden, door bijvoorbeeld pathogenen of onderlinge concurrentie. Hierdoor ontstaat een vicieuze cirkel; de verzwakte bomen moeten steeds relatief meer energie steken in herstel dan de vitale bomen. De energiereserves van deze bomen worden jaar na jaar verder uitgeput door herstelkosten en productie van zaad, wat resulteert in de geobserveerde dalende groeitrend. Door het beschadigde wortelstelsel waaien de bomen ook gemakkelijker om tijdens een storm (Campioli et al., 2012; DeSoto et al., 2020; Dulamsuren et al., 2022; Kint Vincentand Aertsen, 2012; Leuschner, 2020).
De nog levende beuken herstelden zeer snel na een droogte (Figuur 4). Ook werd er bij de nog levende bomen slechts een zeer geringe algemene groeidaling gevonden (Figuur 3). Deze bevindingen zijn gelijkaardig aan een studie van beuken uit het Meerdaalwoud en Zoniënwoud door Vannoppen et al. (2018). Dat beuken in het Zoniënwoud het relatief vaak beter doen onder het veranderend klimaat in vergelijking met andere bossen is volgens ons te wijten aan twee factoren. Ten eerste is de bodem op deze site van uitzonderlijk hoge kwaliteit voor beuk: de diepe leembodem kan veel water opslaan dat vervolgens gebruikt kan worden tijdens een droogte. Ten tweede ligt dit bestand centraal in een groot boscomplex. Hierdoor wordt hitte en droogte tijdens hittegolven sterker gebufferd dan in kleine boscomplexen of aan de bosrand (De Frenne et al., 2021). Deze oude beuken doen het tot nu toe dus nog redelijk goed en zijn voorlopig behoorlijk opgewassen tegen de negatieve effecten van klimaatverandering.
In de toekomst kan een verdere uitval van droogtegevoelige individuen verwacht worden. Onze resultaten suggereren echter dat beuk niet snel uit het Zoniënwoud zal verdwijnen, ook al zullen deze bomen vermoedelijk een verminderde groei vertonen naarmate droogte en hittegolven steeds vaker voorkomen.
Tot slot
Tot slot willen we de geïnteresseerde lezer graag verwijzen naar het wetenschappelijk artikel dat de basis vormt voor dit Bosrevue artikel (https://doi.org/10.1016/j.scitotenv.2023.166148) en naar een artikel over dit onderzoek dat in de Standaard verscheen (https://www.standaard.be/cnt/dmf20230821_96026109).
Ook wil ik (Louis) graag deze kans nemen om prof. dr. ir. Jan Van den Bulcke, prof. dr. ir. Tom De Mil, dr. ir. Kristof Haneca en dr. ir. Kris Vandekerkhove te bedanken voor hun begeleiding tijdens mijn thesis (waarop dit onderzoek gebaseerd is). Ook wil ik het hele boorteam bedanken voor de geleverde inspanningen, merci Jaime, Stijn, Toon, Laura en Ilié. Mijn dank gaat ook uit aan het ANB en het INBO voor jullie ondersteuning en toelating voor de staalnames.
Gelieve als volgt te citeren:
Louis Verschuren, Tom De Mil, Kristof Haneca, Kris Vandekerkhove, Pieter De Frenne, Joris Van Acker, Jan Van den Bulcke (2023) Heading for a fall? Het groeipatroon van levende en windgeworpen beuken vertelt ons meer. Bosrevue 109a
ISSN 2565-6953 – Bosrevue 109a
-
Referenties
Allen, C. D., Breshears, D. D., & McDowell, N. G. (2015). On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere, 6(8), art129.
Baeté, H., De Keersmaeker, L., Van de Kerckhove, P., Christiaens, B., Esprit, M., & Vandekerkhove, K. (2002). Bosreservaat Kersselaerspleyn: basisrapport: algemene situering, standplaatsbeschrijving, historische kadering en overzicht: wetenschappelijk onderzoek (Issue 5). Instituut voor Bosbouw en Wildbeheer.
Brinkmann, N., Eugster, W., Buchmann, N., & Kahmen, A. (2019). Species-specific differences in water uptake depth of mature temperate trees vary with water availability in the soil. Plant Biology, 21(1), 71–81.
Campioli, M., Vincke, C., Jonard, M., Kint, V., Demarée, G., & Ponette, Q. (2012). Current status and predicted impact of climate change on forest production and biogeochemistry in the temperate oceanic European zone: review and prospects for Belgium as a case study. Journal of Forest Research, 17(1), 1–18.
Cavin, L., & Jump, A. S. (2017). Highest drought sensitivity and lowest resistance to growth suppression are found in the range core of the tree Fagus sylvatica L. not the equatorial range edge. Global Change Biology, 23(1), 362–379.
De Frenne, P., Lenoir, J., Luoto, M., Scheffers, B. R., Zellweger, F., Aalto, J., Ashcroft, M. B., Christiansen, D. M., Decocq, G., De Pauw, K., Govaert, S., Greiser, C., Gril, E., Hampe, A., Jucker, T., Klinges, D. H., Koelemeijer, I. A., Lembrechts, J. J., Marrec, R., … Hylander, K. (2021). Forest microclimates and climate change: Importance, drivers and future research agenda. Global Change Biology, 27(11), 2279–2297. https://doi.org/10.1111/GCB.15569
De Mil, T., Vannoppen, A., Beeckman, H., Van Acker, J., & Van den Bulcke, J. (2016). A field-to-desktop toolchain for X-ray CT densitometry enables tree ring analysis. Annals of Botany, 117, mcw063.
Demol, M., Vandekerkhove, K., De Keersmaeker, L., & Van de Kerckhove, P. (2020). Het Zoniënwoud in 3D opmeten: op stap met een laserscanner. Bosreservatennieuws , 17, 26–30.
Van den Bulcke, J., Boone, M., Dhaene, J., Denis, V., Van Hoorebeke, L., Boone, M., Wyffels, F., Beeckman, H., Van Acker, J., & De Mil, T. (2019). Advanced X-ray CT scanning can boost tree-ring research for earth-system sciences. Annals of Botany, 124, 837–847.
DeSoto, L., Cailleret, M., Sterck, F., Jansen, S., Kramer, K., Robert, E. M. R., Aakala, T., Amoroso, M. M., Bigler, C., Camarero, J. J., Čufar, K., Gea-Izquierdo, G., Gillner, S., Haavik, L. J., Hereş, A. M., Kane, J. M., Kharuk, V. I., Kitzberger, T., Klein, T., … Martínez-Vilalta, J. (2020). Low growth resilience to drought is related to future mortality risk in trees. Nature Communications, 11(1). https://doi.org/10.1038/s41467-020-14300-5
Dittmar, C., Zech, W., & Elling, W. (2003). Growth variations of Common beech (Fagus sylvatica L.) under different climatic and environmental conditions in Europe—a dendroecological study. Forest Ecology and Management, 173(1), 63–78.
Drobyshev, I., Övergaard, R., Saygin, I., Niklasson, M., Hickler, T., Karlsson, M., & Sykes, M. T. (2010). Masting behaviour and dendrochronology of European beech (Fagus sylvatica L.) in southern Sweden. Forest Ecology and Management, 259(11), 2160–2171.
Dulamsuren, C., Bat-Enerel, B., Meyer, P., & Leuschner, C. (2022). Did stand opening 60 years ago predispose a European beech population to death? Trees, Forests and People, 8, 100265.
Dyderski, M. K., Paź, S., Frelich, L. E., & Jagodziński, A. M. (2018). How much does climate change threaten European forest tree species distributions? Global Change Biology, 24(3), 1150–1163. https://doi.org/10.1111/GCB.13925
Giagli, K., Gričar, J., Vavrč\’\ik, H., Menš\’\ik, L., & Gryc, V. (2016). The effects of drought on wood formation in Fagus sylvatica during two contrasting years. IAWA Journal, 37(2), 332–348.
Goris, R. (2014). Het grondvlak. Bosrevue, 20, 17.
Hacket-Pain, A. J., Friend, A. D., Lageard, J. G. A., & Thomas, P. A. (2015). The influence of masting phenomenon on growth–climate relationships in trees: explaining the influence of previous summers’ climate on ring width. Tree Physiology, 35(3), 319–330.
Haneca, K. (2017). Dendrochronologie en erfgoedonderzoek (Issue 16). Agentschap Onroerend Erfgoed.
Kint Vincent, Aertsen, W., C. M., V. D., D. A., M. B. (2012). Radial growth change of temperate tree species in response to altered regional climate and air quality in the period 1901–2008. Climatic Change, 115(2), 343–363.
Köcher, P., Gebauer, T., Horna, V., & Leuschner, C. (2009). Leaf water status and stem xylem flux in relation to soil drought in five temperate broad-leaved tree species with contrasting water use strategies. Annals of Forest Science, 66(1), 1.
Latte, N. (2017). Comment le changement climatique influence-t-il la croissance du hêtre (Fagus sylvatica L.) le long du gradient bioclimatique de la Belgique? Une approche dendroécologique. Université de Liège, Liège, Belgique.
Latte, N., & Claessens, H. (2015). Analyse de l’influence du changement climatique sur la croissance du hêtre en Forêt Soignes.
Latte, N., Kint, V., Drouet, T., Penninckx, V., Lebourgeois, F., Vanwijnsberghe, S., & Claessens, H. (2015). Dendroécologie du hêtre en forêt de Soignes : Les cernes des arbres nous renseignent sur les changements récents et futurs. Forêt.Nature, 137(Octobre-Novembre-Décembre), 24–37.
Laubhann, D., Sterba, H., Reinds, G. J., & De Vries, W. (2009). The impact of atmospheric deposition and climate on forest growth in European monitoring plots: An individual tree growth model. Forest Ecology and Management, 258(8), 1751–1761.
Leuschner, C. (2020). Drought response of European beech (Fagus sylvatica L.)—A review. Perspectives in Plant Ecology, Evolution and Systematics, 47, 125576.
Leuschner, C., Wedde, P., L. T. (2019). The relation between pressure–volume curve traits and stomatal regulation of water potential in five temperate broadleaf tree species. Annals of Forest Science, 76(2), 60.
Martinez del Castillo, E., Zang, C. S., Buras, A., Hacket-Pain, A., Esper, J., Serrano-Notivoli, R., Hartl, C., Weigel, R., Klesse, S., Resco de Dios, V., Scharnweber, T., Dorado-Liñán, I., van der Maaten-Theunissen, M., van der Maaten, E., Jump, A., Mikac, S., Banzragch, B. E., Beck, W., Cavin, L., … de Luis, M. (2022). Climate-change-driven growth decline of European beech forests. Communications Biology, 5(1). https://doi.org/10.1038/s42003-022-03107-3
Müller-Haubold Hilmar, Hertel, D., L. C. (2015). Climatic Drivers of Mast Fruiting in European Beech and Resulting C and N Allocation Shifts. Ecosystems, 18(6), 1083–1100.
Müller-Haubold Hilmar, Hertel, D., S. D., K. F., L. C. (2013). Climate Responses of Aboveground Productivity and Allocation in Fagus sylvatica: A Transect Study in Mature Forests. Ecosystems, 16(8), 1498–1516.
Reyer, C. P. O., Leuzinger, S., Rammig, A., Wolf, A., Bartholomeus, R. P., Bonfante, A., de Lorenzi, F., Dury, M., Gloning, P., Abou Jaoudé, R., Klein, T., Kuster, T. M., Martins, M., Niedrist, G., Riccardi, M., Wohlfahrt, G., de Angelis, P., de Dato, G., François, L., … Pereira, M. (2013). A plant’s perspective of extremes: terrestrial plant responses to changing climatic variability. Global Change Biology, 19(1), 75–89.
Scherrer, D., Bader, M. K.-F., & Körner, C. (2011). Drought-sensitivity ranking of deciduous tree species based on thermal imaging of forest canopies. Agricultural and Forest Meteorology, 151(12), 1632–1640.
Speer, J. (2010). Fundamentals of Tree Ring Research (pp. 19–47). University of Arizona Press.
Termonia, P., Van Schaeybroeck, B., De Cruz, L., De Troch, R., Caluwaerts, S., Giot, O., Hamdi, R., Vannitsem, S., Duchêne, F., Willems, P., Tabari, H., Van Uytven, E., Hosseinzadehtalaei, P., Van Lipzig, N., Wouters, H., Vanden Broucke, S., van Ypersele, J.-P., Marbaix, P., Villanueva-Birriel, C., … Pottiaux, E. (2018). The CORDEX.be initiative as a foundation for climate services in Belgium. Climate Services, 11, 49–61.
Tsen, E. W. J., Sitzia, T., & Webber, B. L. (2016). To core, or not to core: the impact of coring on tree health and a best-practice framework for collecting dendrochronological information from living trees. Biological Reviews, 91(4), 899–924. https://doi.org/10.1111/brv.12200
Vacchiano, G., Hacket-Pain, A., Turco, M., Motta, R., Maringer, J., Conedera, M., Drobyshev, I., & Ascoli, D. (2017). Spatial patterns and broad-scale weather cues of beech mast seeding in Europe. New Phytologist, 215(2), 595–608.
Vandekerkhove, K., Vanhellemont, M., Vrška, T., Meyer, P., Tabaku, V., Thomaes, A., Leyman, A., De Keersmaeker, L., & Verheyen, K. (2018). Very large trees in a lowland old-growth beech (Fagus sylvatica L.) forest: Density, size, growth and spatial patterns in comparison to reference sites in Europe. Forest Ecology and Management, 417, 1–17.
Van den Bulcke, J., Wernersson, E. L. G., Dierick, M., Van Loo, D., Masschaele, B., Brabant, L., Boone, M. N., Van Hoorebeke, L., Haneca, K., Brun, A., Luengo Hendriks, C. L., & Van Acker, J. (2014). 3D tree-ring analysis using helical X-ray tomography. Dendrochronologia, 32(1), 39–46. https://doi.org/10.1016/j.dendro.2013.07.001
van der Werf, G. W., Sass-Klaassen, U. G. W., & Mohren, G. M. J. (2007). The impact of the 2003 summer drought on the intra-annual growth pattern of beech (Fagus sylvatica L.) and oak (Quercus robur L.) on a dry site in the Netherlands. Dendrochronologia, 25(2), 103–112.
Vanhellemont, M., Sousa-Silva, R., Maes, S. L., Van den Bulcke, J., Hertzog, L., De Groote, S. R. E., Van Acker, J., Bonte, D., Martel, A., Lens, L., & Verheyen, K. (2019). Distinct growth responses to drought for oak and beech in temperate mixed forests. Science of the Total Environment, 650, 3017–3026. https://doi.org/10.1016/j.scitotenv.2018.10.054
Vannoppen, A., Boeckx, P., De Mil, T., Kint, V., Ponette, Q., Van den Bulcke, J., Verheyen, K., & Muys, B. (2018). Climate driven trends in tree biomass increment show asynchronous dependence on tree-ring width and wood density variation. Dendrochronologia, 48, 40–51.


