


Bosrevue

Samenvatting
Grote solitaire bomen zijn zowel in stedelijke als landelijke gebieden belangrijk voor biodiversiteit. Helaas zijn deze bomen bijzonder gevoelig voor klimaatverandering en worden ze wereldwijd zeldzamer door intensief landgebruik. Momenteel wordt er weinig aandacht besteed aan het specifieke microklimaat gecreëerd door solitaire bomen. In dit artikel bekijken we hoe verstedelijking en boomsoort (zomereik, linde en es) de luchttemperatuur onder de kroon van solitaire bomen beïnvloedt. Uit ons onderzoek blijkt dat de temperatuur onder de kroon van bomen in stedelijke gebieden warmer is dan onder bomen in landelijke gebieden: 10% extra verstedelijking, bepaald aan de hand van de hoeveelheid bebouwde oppervlakte, zorgde voor een 0,10°C hogere gemiddelde zomertemperatuur. Zomereik en linde hadden een grotere invloed op de luchttemperatuur dan es: ze verlaagden de maximumtemperatuur onder hun kroon 0,55°C meer dan es. Dit toont aan dat de boomsoort een impact heeft op het microklimaat en benadrukt het belang van een bewuste soortkeuze in steden.
Inleiding
Grote solitaire bomen zijn alleenstaande bomen die geen directe interactie hebben met andere bomen in hun omgeving. Ze hebben vaak een indrukwekkende vorm omdat ze geen concurrentie van andere bomen ervaren en daardoor een brede kroon kunnen ontwikkelen. Grote solitaire bomen zijn belangrijke landschapselementen met een hoge culturele en ecologische waarde (Manning et al., 2006). Ze vormen een habitat voor allerlei soorten insecten, vogels en zoogdieren, verhogen de plantenrijkdom en hebben een disproportioneel grote invloed op het lokale ecosysteem, bijvoorbeeld via microklimaatbuffering (Fischer et al., 2010). Helaas verdwijnen ze wereldwijd door intensief landgebruik en de effecten van klimaatverandering. Solitaire bomen zijn namelijk gevoeliger voor een veranderend klimaat omdat ze niet beschermd worden door de bufferende werking van andere bomen in hun omgeving, zoals wel het geval is in een bos (Arumugam et al., 2024; Lindenmayer & Laurance, 2017).
Microklimaten zijn lokale variaties op het grootschalige macroklimaat. In de zomer is het onder bomen doorgaans koeler, terwijl het in de winter onder bomen net warmer is. In regio’s met een vlak reliëf wordt het microklimaat onder een boom vooral gecreëerd door schaduw en door transpiratie door de boom. De morfologische kenmerken van boomsoorten spelen hierbij een rol (Rahman et al., 2020; Winbourne et al., 2020). Bomen transpireren, waardoor ze de omgeving afkoelen en de luchtvochtigheid verhogen. Door inkomende zonnestraling te blokkeren, voorkomen bomen ook de absorptie van kortgolvige straling in oppervlaktematerialen, en de daaropvolgende emissie van langgolvige warmtestraling die de omgeving opwarmt. Zowel het schaduwwerpend vermogen als de transpiratie van bomen zijn afhankelijk van de kroondichtheid of LAI (leaf area index, hoeveelheid bladoppervlak per m²). Boomsoorten met een hoge LAI zoals beuk en linde hebben een dichte kroon die meer schaduw werpt en daardoor de omgeving meer kan afkoelen. Soorten zoals populier en es met een lage LAI hebben een meer open kroon en werpen dus minder schaduw (Rahman et al., 2019). Het microklimaat is een combinatie van temperatuur en luchtvochtigheid, maar in dit artikel werd ervoor gekozen om enkel het temperatuur effect te belichten. In het Engelstalige wetenschappelijk artikel waarop dit artikel gebaseerd is (Van Den Bossche et al., 2025; Microclimate of large solitary trees | sciencedirect.com), kunnen ook resultaten in verband met luchtvochtigheid worden teruggevonden.
Stedelijke omgevingen worden gekenmerkt door het stedelijk hitte-eilandeffect, waarbij temperaturen in de stedelijke omgeving hoger zijn dan die in het omringende landelijk gebied. Dit effect wordt veroorzaakt door de hoge hoeveelheid niet-natuurlijke materialen die overdag veel warmte kunnen opslaan (vb. asfalt, beton), de straatarchitectuur, en warmteproductie door onder andere verkeer en airco’s (Ward et al., 2016). Stedelijke groenelementen zoals parken hebben een koelend effect, waardoor ze de sterkte van het stedelijk hitte-eiland verminderen en inwoners, fauna en flora beschermen tegen extreme hitte (Wong et al., 2021). Meer dan 70% van de Europese bevolking woont in verstedelijkt gebied (dorpen en voorsteden: 36,4%, steden: 39,1%). Het mitigeren van stedelijk hitte-eilanden is dan ook erg belangrijk voor de mens maar ook voor plant en dier (Grimm et al., 2008).
Grootschalige kwantitatieve studies die het microklimaat langsheen een verstedelijkingsgradiënt onderzoeken, ontbreken. Microklimaatonderzoek naar solitaire bomen betreft voornamelijk lokale casestudies, is gericht op een specifiek ecosysteem (bv. savannes) of focust op andere aspecten die het microklimaat kunnen beïnvloeden zoals transpiratie. Wij hebben het microklimaat gemeten onder de kroon van solitaire bomen langsheen een verstedelijkingsgradiënt, bepaald aan de hand van de hoeveelheid bebouwde oppervlakte, in en rond negen Europese steden. We bestudeerden drie boomsoorten met contrasterende krooneigenschappen, namelijk zomereik (Quercus robur), es (Fraxinus excelsior) en linde (Tilia sp.). Het doel van ons onderzoek was inzicht krijgen in hoe verstedelijking het microklimaat van grote solitaire bomen beïnvloedt en welke rol boomsoort hierin speelt. Dit kadert in een breder onderzoek naar de biodiversiteit die gerelateerd is aan grote solitaire bomen (CoolTree project).
Methodes
We verzamelden microklimaatdata onder de kroon van 216 grote solitaire bomen in negen Europese steden langsheen een latitudinale gradiënt van Noord-Frankrijk tot Centraal-Noorwegen, en een longitudinale gradiënt van Noord-Frankrijk tot Zuid-Polen (Fig. 1). Op deze manier omvat ons onderzoek data uit verschillende klimatologische regio’s.
In elke stad bestudeerden we acht zomereiken, acht lindes en acht essen, elk langsheen een verstedelijkingsgradiënt (een gradiënt per bestudeerde boomsoort). Zomereik komt vaak solitair voor, is relatief droogtetolerant, en heeft een wijde en halfopen kroon, vaak met zware zijtakken. Linde is de meest voorkomende boom in stedelijke gebieden in Europa, is droogte- en warmtetolerant, en heeft een dichte kroon die veel schaduw werpt. Es komt tevens vaak voor als grote solitaire boom en verkiest vochtigere bodems maar kan droogtetolerant zijn wanneer de bodem voldoende kalk bevat. De soort heeft een grote, open kroon. De laatste decennia heeft es te maken met een invasieve schimmelziekte die essentaksterfte veroorzaakt. Deze ziekte wordt gekenmerkt door het afsterven van bladeren en takken, en kan na enkele jaren resulteren in de dood van de aangetaste boom (Pautasso et al., 2013). Wij bekeken enkel essen die geen of weinig symptomen van essentaksterfte vertoonden.
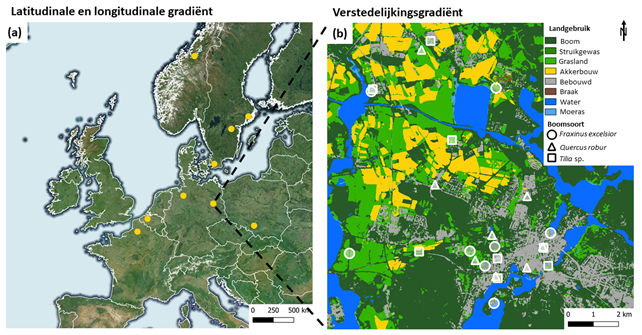
Figuur 1: Overzicht van de studielocaties met (a) de negen onderzochte steden van zuid naar noord: Amiens, Tarnowskie Góry, Gent, Potsdam, Bremen, Lund, Linköping, Stockholm, en Trondheim; en (b) de onderzochte grote solitaire bomen langsheen de verstedelijkingsgradiënt in Potsdam (Oost-Duitsland) met als achtergrond het lokale landgebruik.
We selecteerden onze studiebomen op basis van hun grootte, solitair karakter, en de verstedelijkingsgraad van de omgeving. De gemiddelde stamdiameter was 94 cm. De dunste boom had een stamdiameter van 26 cm, de grootste had een stamdiameter van 244 cm. De boomkronen hadden minimale interactie met naburige bomen en hebben zich vrij kunnen ontwikkelen. We bepaalden de verstedelijking in een straal van 75 m rond de geselecteerde bomen aan de hand van de hoeveelheid bebouwd oppervlak. Het percentage verstelijking varieerde van 0% (open, landelijke omgeving) tot 98% (stadscentrum). We onderzochten enkel bomen met een boomspiegel van minstens 100m².
We brachten het microklimaat in kaart met behulp van sensoren die de luchttemperatuur elk half uur registreerden gedurende tien maanden (begin juli 2022 tot eind maart 2023). De sensoren hingen aan de noordkant van de bomen op ongeveer drie meter hoogte tegen de stam, en werden beschermd tegen neerslag en zonlicht door witte pvc-buizen. Van de 216 gebruikte sensoren leverden 19 geen data op omwille van defecten of vandalisme.
We combineerden de temperatuurmetingen tot maandelijkse gemiddelden voor elke boom (maandelijks minimum, maximum en gemiddelde). We berekenden voor elke boom het verschil (ΔT) tussen het maandelijks gemiddelde onder de boom zelf en het maandelijks gemiddelde voor alle bomen in de stad (zie Box 1). Een positief verschil betekent dat het onder de kroon van de solitaire boom warmer was dan gemiddeld onder solitaire boomkronen in de stad, een negatief verschil betekende dat het onder de boomkroon koeler was dan gemiddeld. Vervolgens onderzochten we de impact van verstedelijking, boomsoort en stamdiameter op de gemiddelde maandelijkse temperatuur (minimum, maximum, gemiddelde).
Box 1 – statistische analyses
Metingen: We gebruikten Lascar-EL-USB2 sensoren om elk half uur de luchttemperatuur en luchtvochtigheid onder de kroon van 216 grote solitaire bomen te meten.
Dagelijkse waarden: De half-uurlijkse temperatuurmetingen werden opgeschoond en onderworpen aan een visuele kwaliteitscheck, waarbij onrealistische meetpunten verwijderd werden. Vervolgens werd deze opgeschoonde data gebruikt om de dagelijkse minimum, maximum en gemiddelde temperatuur te berekenen.
Maandelijkse waarden: De dagelijkse waarden werden gecombineerd tot maandelijkse waarden door per maand het gemiddelde te berekenen van de dagelijkse minimum, maximum en gemiddelde temperatuur.
Modelopbouw: We gebruikten linear mixed effects models met de volgende structuur:

Deze modellen gebruiken het verschil tussen de maandelijkse minimum, maximum of gemiddelde temperatuur onder de boom en het maandelijks minimum, maximum of gemiddelde voor alle bomen in de stad (ΔT) als responsvariabele. Verstedelijking, boomsoort, stamdiameter en het maandelijks minimum, maximum of gemiddelde voor alle bomen in de stad zijn verklarende variabelen.
We bekeken het resultaat van deze modellen wanneer we alle data in acht namen (juli 2022 – maart 2023), maar onderzochten ook de resultaten wanneer we enkel data van de zomermaanden (juli 2022 en augustus 2022) en enkel de wintermaanden (december 2022 en januari 2023) gebruikten.
Resultaten
In de zomer (juli – augustus) was de minimumtemperatuur (maandgemiddelde) onder solitaire bomen in stedelijke omgeving (>70% bebouwde oppervlakte rond de boom) gemiddeld 1,44°C hoger dan onder solitaire bomen in landelijke omgeving (<10% bebouwde oppervlakte rond de boom). Ook de gemiddelde zomertemperatuur (1,14°C hoger) en de maximale zomertemperatuur (0,74°C) waren hoger onder de kroon van bomen in sterk verstedelijkte omgeving.
Voor de hele studieperiode en het hele studiegebied, steeg het verschil tussen de gemiddelde maandelijkse minimumtemperatuur en de gemiddelde minimumtemperatuur in de stad (ΔTmin) 0,10°C wanneer de verstedelijking in een straal van 75 m rond de boom met 10% toenam. Het verschil tussen de gemiddelde temperatuur en de gemiddelde temperatuur in de stad (ΔTgem) steeg 0,06°C per 10% extra verstedelijking. De invloed van verstedelijking was groter in de zomer dan in de winter. ΔTmin steeg in de zomer 0,17°C wanneer de verstedelijking met 10% toenam, terwijl deze in de winter slechts 0,07°C toenam. ΔTgem werd minder beïnvloed, maar steeg bij toenemende verstedelijking nog steeds meer in de zomer dan in de winter. We vonden geen significant effect van verstedelijking op ΔTmax (Fig. 2).

Figuur 2: De afwijking van de (a) minimum, (b) gemiddelde, en (c) maximum luchttemperatuur onder de kroon van een solitaire boom ten opzichte van alle onderzochte bomen in de stad voor elke dag in de studieperiode. De kleur van de lijn toont het percentage verstedelijking in een straal van 75 m rondom de boom (rood: bijna volledig bebouwd; blauw: veel open ruimte). Een negatieve temperatuurafwijking betekent dat de temperatuur onder de boom lager is dan de gemiddelde temperatuur onder alle bomen in de stad. Een positieve temperatuurafwijking betekent dat de temperatuur onder de boom hoger is.
Onder zomereik was het koeler dan onder es: de gemiddelde maandtemperatuur was 0,12°C lager, de maximummaandtemperatuur 0,23°C lager (voor het hele studiegebied en de hele studieperiode). Onder zomereik was de gemiddelde maandtemperatuur ook lager dan onder linde (0,11°C), maar de maximumtemperatuur verschilde niet (Fig. 3). Voor de hele studieperiode en het hele studiegebied vonden we geen temperatuurverschillen tussen linde en es. De verschillen tussen de boomsoorten waren groter in de zomer dan in de winter (Fig. 3). De maximumtemperatuur in de zomer was onder es 0,51°C hoger dan onder zomereik en 0,58°C hoger dan onder linde. De gemiddelde temperatuur was 0,21°C hoger onder es dan onder zomereik en 0,19°C hoger dan onder linde. Ook in de winter vonden we een effect van boomsoort: de gemiddelde temperatuur onder linde was 0,19°C hoger dan onder zomereik. Minimumtemperaturen verschilden niet tussen de drie boomsoorten (niet in de hele periode, in de winter, of in de zomer).
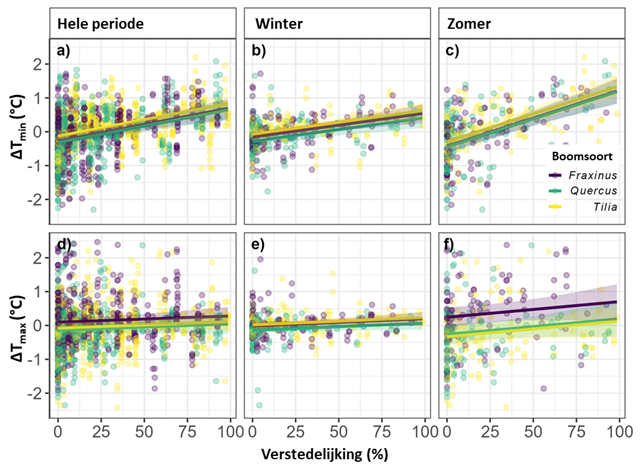
Figuur 3: De afwijking van de gemiddelde minimummaandtemperatuur (a, b, c,) en maximummaandtemperatuur (d, e, f) ten opzichte van het gemiddelde voor alle 24 bomen in de stad in relatie tot de verstedelijking in een straal van 75 m rond de solitaire boom voor (a, d) de hele studieperiode, (b, e) winter, en (c, f) zomer. Een negatieve temperatuurafwijking betekent dat de temperatuur onder de boom lager is dan de gemiddelde temperatuur onder alle bomen in de stad. Een positieve temperatuurafwijking betekent dat de temperatuur onder de boom hoger is. De grafiek toont temperatuurvoorspellingen samen met het 95%-betrouwbaarheidsinterval per boomsoort.
Discussie
Onze resultaten tonen aan dat de temperatuur onder de kroon van grote solitaire bomen hoger was in stedelijke gebieden dan in landelijke gebieden, en dat verschillen langsheen de verstedelijkingsgradiënt en tussen boomsoorten het grootst waren in de zomer.
Steden bevatten veel verharde oppervlakken die straling en warmte kunnen vasthouden, waardoor de temperatuur in stedelijke omgevingen vaak warmer is. Wij vonden dat verstedelijking ook leidde tot het stijgen van de temperatuur gemeten onder de kroon van grote solitaire bomen. Dit is vergelijkbaar met resultaten uit voorgaande studies, waar bijvoorbeeld de maximumzomertemperatuur 0,15°C steeg wanneer de verstedelijking in een straal van 500 m rond het meetpunt met 10% toenam (Schatz & Kucharik, 2014). Onze resultaten lieten een kleiner effect van verstedelijking zien. Het regionale macroklimaat en de stedelijke context kunnen een belangrijke invloed hebben op de grootte van het stedelijk hitte-eilandeffect, en dus ook op het effect van verstedelijking (Zhang et al., 2022). Deze afhankelijkheid kan verklaren waarom effectgroottes verschillend zijn. Bomen waarbij de hoeveelheid verharde oppervlakte in een straal van 75 m laag is, kunnen nog steeds in sterk verstedelijkt gebied groeien, zoals vaak het geval is voor parkbomen.
De effecten van verstedelijking waren het grootst op de minimumtemperaturen in de zomer. Het stedelijk hitte-eilandeffect is het grootst gedurende de nacht, wanneer luchttemperaturen laag zijn en steen, asfalt en beton de opgeslagen warmte afgeven, en heeft op die manier een belangrijke invloed op minimumtemperaturen. In stedelijke omgevingen ligt de minimumzomertemperatuur inmiddels meer dan 2°C boven het niveau van de referentieperiode 1850-1900 (VMM, n.d). Onder stedelijke bomen blijkt de minimumzomertemperatuur gemiddeld zelfs al 1,44°C hoger te zijn dan onder bomen in landelijk gebied. Deze aanhoudende opwarming kan verstrekkende gevolgen hebben voor de menselijke gezondheid (Gillerot et al., 2024) en beïnvloedt eveneens de fenologie en voortplanting van planten en dieren en de soortensamenstelling van ecologische gemeenschappen (Nunez et al., 2019).
Verschillende boomsoorten hebben uiteenlopende effecten op temperatuurmitigatie. Zo verlaagden linde en eik de maximale zomertemperaturen met 0,5°C meer dan es. Dit verschil is waarschijnlijk te verklaren door de hogere LAI en het groter schaduwwerpend vermogen van linde en eik (Sjöman et al., 2021; Verheyen et al., 2012). Hoewel we essen selecteerden met weinig symptomen van essentaksterfte, is het mogelijk dat de ziekte het soortverschil versterkte en het temperatuurverlagende effect van es verminderde.
In ons onderzoek vonden we dat gemiddelde wintertemperaturen onder linde 0,19°C hoger waren dan onder eik, met es tussenin. Ook in de winter blijven bomen warmtestraling onderscheppen. Bomen zonder bladeren kunnen tot 50% van de inkomende zonnestraling absorberen, afhankelijk van hun kroonstructuur (Wong et al., 2021). De verschillen die wij vaststelden zijn dus mogelijks deels toe te schrijven aan de specifieke kroonstructuur van de onderzochte bomen.
Conclusie
Deze studie benadrukt de rol van grote solitaire bomen in stedelijke en landelijke omgevingen. Het onderzoek wijst daarbij op het belang van het beschermen van deze bomen, zowel in stedelijk als landelijk gebied. Er is behoefte aan geïntegreerde bomenplannen die verder gaan dan de huidige beschermingsmaatregelen en niet enkel inzetten op de bescherming van aanwezige bomen, maar ook ruimte creëren voor nieuwe, jonge bomen om uit te groeien tot grote, oude exemplaren. Daarnaast is het essentieel dat er voldoende ruimte en onverharde oppervlakten onder de kroon van deze bomen wordt voorzien om de gezondheid van de bomen te vrijwaren en sterfte te minimaliseren. De biodiversiteit en ecosysteemdiensten die grote bomen leveren, kunnen niet zomaar worden vervangen door die van jonge bomen. Daarom is een toekomstgerichte aanpak nodig waarin zowel het behoud van oude bomen als de groei van nieuwe bomen centraal staat.
Het koelend effect van solitaire bomen hangt af van verschillende factoren, waaronder de boomsoort. Hierom adviseren wij stadsplanners en groenbeheerders om bij de selectie van boomsoorten niet alleen esthetische aspecten, klimaatrobuustheid, groeisnelheid en allergeenpotentie te overwegen, maar ook de invloed van de soort op het lokale microklimaat. Tegelijkertijd is verder onderzoek nodig naar de rol van boomsoortkeuze, kroonstructuur en beheergewoonten zoals snoeien, om de kennis over microklimatologische effecten van solitaire bomen te vergroten.
Door grote solitaire bomen te erkennen als kerncomponenten van stedelijk groen, kunnen steden en gemeenten bijdragen aan een gunstig microklimaat, biodiversiteit ondersteunen, en op die manier een meer leefbare omgeving creëren. Het beschermen van huidige en toekomstige solitaire bomen sluit aan bij de doelstellingen van de Europese Green Deal, en draagt bij aan het streven van Europa om tegen 2030 drie miljard bomen te planten (Trees for Life; European Committee of the Regions, 2022).
Dankwoord
Met dank aan het Fonds voor Wetenschappelijk Onderzoek (FWO) voor het financieren van het CoolTree-project (no. G000322N) en het wetenschappelijk onderzoeksnetwerk FLEUR (https://fleur.ugent.be/).
Gelieve als volgt te citeren:
Astrid Van den Bossche, Karlien Moeys, Karen De Pauw, Koenraad Van Meerbeek, Arno Thomaes, Thomas Vanneste, Louis Verschuren, Pieter De Frenne (2025) Het microklimaat van grote solitaire bomen. Bosrevue 120a.
ISSN 2565-6953 – Bosrevue 120a
-
Referenties
Arumugam, R., Guichard, F., & Lutscher, F. (2024). Early warning indicators capture catastrophic transitions driven by explicit rates of environmental change. Ecology. https://doi.org/10.1002/ecy.4240
European Committee of the Regions. (2022). Trees for life. Https://Cor.Europa.Eu/En/Engage/Pages/Trees-for-Life.Aspx.
Fischer, J., Stott, J., & Law, B. S. (2010). The disproportionate value of scattered trees. Biological Conservation, 143(6), 1564–1567. https://doi.org/10.1016/j.biocon.2010.03.030
Gillerot, L., Landuyt, D., De Frenne, P., Muys, B., & Verheyen, K. (2024). Urban tree canopies drive human heat stress mitigation. Urban Forestry and Urban Greening, 92. https://doi.org/10.1016/j.ufug.2023.128192
Grimm, N. B., Faeth, S. H., Golubiewski, N. E., Redman, C. L., Wu, J., Bai, X., & Briggs, J. M. (2008). Global change and the ecology of cities. Science, 319(5864), 756–760. https://doi.org/10.1126/science.1150195
Lindenmayer, D. B., & Laurance, W. F. (2017). The ecology, distribution, conservation and management of large old trees. Biological Reviews, 92(3), 1434–1458. https://doi.org/10.1111/brv.12290
Manning, A. D., Fischer, J., & Lindenmayer, D. B. (2006). Scattered trees are keystone structures – Implications for conservation. Biological Conservation, 132(3), 311–321. https://doi.org/10.1016/j.biocon.2006.04.023
Nunez, S., Arets, E., Alkemade, R., Verwer, C., & Leemans, R. (2019). Assessing the impacts of climate change on biodiversity: is below 2 °C enough? Climatic Change, 154(3–4), 351–365. https://doi.org/10.1007/s10584-019-02420-x
Pautasso, M., Aas, G., Queloz, V., & Holdenrieder, O. (2013). European ash (Fraxinus excelsior) dieback – A conservation biology challenge. In Biological Conservation (Vol. 158, pp. 37–49). https://doi.org/10.1016/j.biocon.2012.08.026
Rahman, M. A., Moser, A., Rötzer, T., & Pauleit, S. (2019). Comparing the transpirational and shading effects of two contrasting urban tree species. Urban Ecosystems, 22(4), 683–697. https://doi.org/10.1007/s11252-019-00853-x
Rahman, M. A., Stratopoulos, L. M. F., Moser-Reischl, A., Zölch, T., Häberle, K. H., Rötzer, T., Pretzsch, H., & Pauleit, S. (2020). Traits of trees for cooling urban heat islands: A meta-analysis. In Building and Environment (Vol. 170). Elsevier Ltd. https://doi.org/10.1016/j.buildenv.2019.106606
Schatz, J., & Kucharik, C. J. (2014). Seasonality of the urban heat island effect in Madison, Wisconsin. Journal of Applied Meteorology and Climatology, 53(10), 2371–2386. https://doi.org/10.1175/JAMC-D-14-0107.1
Sjöman, J., Hirons, A., Bassuk, N., & Sjöman, H. (2021). Plant and wood area index of solitary trees for urban contexts in nordic cities. Arboriculture & Urban Forestry, 47(6), 252–266. https://doi.org/10.48044/jauf.2021.022
Van den Bossche, A., Moeys, K., De Pauw, K., Van Meerbeek, K., Thomaes, A., Brunet, J., Cousins, S.A.O., Diekmann, M., Graae, B.J., Hagenblad, J., Heavyside, P., Hedwall, Heinken, P. T., …, De Frenne, P. (2025). Microclimate of large solitary trees along rural-to-urban gradients across Europe, Agricultural and Forest Meteorology, Vol. 370, https://doi.org/10.1016/j.agrformet.2025.110585.
Verheyen, K., Baeten, L., De Frenne, P., Bernhardt-Römermann, M., Brunet, J., Cornelis, J., Decocq, G., Dierschke, H., Eriksson, O., Hédl, R., Heinken, T., Hermy, M., Hommel, P., Kirby, K., Naaf, T., Peterken, G., Petřík, P., Pfadenhauer, J., Van Calster, H., … Verstraeten, G. (2012). Driving factors behind the eutrophication signal in understorey plant communities of deciduous temperate forests. Journal of Ecology, 100(2), 352–365. https://doi.org/10.1111/j.1365-2745.2011.01928.x
VMM. (n.d.). Klimaatportaal Vlaanderen – Wat we weten over klimaatverandering. Retrieved December 9, 2024, from https://klimaat.vmm.be/themas/wat-is-klimaatverandering
Ward, K., Lauf, S., Kleinschmit, B., & Endlicher, W. (2016). Heat waves and urban heat islands in Europe: A review of relevant drivers. Science of the Total Environment, 569–570, 527–539. https://doi.org/10.1016/j.scitotenv.2016.06.119
Winbourne, J. B., Jones, T. S., Garvey, S. M., Harrison, J. L., Wang, L., Li, D., Templer, P. H., & Hutyra, L. R. (2020). Tree transpiration and urban temperatures: Current understanding, implications, and future research directions. BioScience, 70(7), 576–588. https://doi.org/10.1093/biosci/biaa055
Wong, N. H., Tan, C. L., Kolokotsa, D. D., & Takebayashi, H. (2021). Greenery as a mitigation and adaptation strategy to urban heat. In Nature Reviews Earth and Environment (Vol. 2, Issue 3, pp. 166–181). Springer Nature. https://doi.org/10.1038/s43017-020-00129-5
Zhang, Z., Paschalis, A., Mijic, A., Meili, N., Manoli, G., van Reeuwijk, M., & Fatichi, S. (2022). A mechanistic assessment of urban heat island intensities and drivers across climates. Urban Climate, 44. https://doi.org/10.1016/j.uclim.2022.101215


