


Bosrevue

Inleiding
Bossen van over de hele wereld zullen geconfronteerd worden met aanzienlijke uitdagingen door klimaatverandering, en dan vooral door een toenemende frequentie en duur van klimaatextremen zoals droogte en hitte (Patacca et al. 2023). Klimaatverandering zal droogteperiodes tijdens het groeiseizoen in West-Europa intensiveren, zoals we in recente jaren reeds gemerkt hebben. Dergelijke klimaatextremen kunnen de vitaliteit van bomen aantasten en zelfs leiden tot boomsterfte, wat op zijn beurt de samenstelling en structuur van bossen kan beïnvloeden (Allen et al. 2010). Om beter te kunnen inschatten wat onze bosecosystemen te wachten staat, is het onder meer van belang om te begrijpen hoe de voorspelde klimaatverandering de reproductie van bomen kan beïnvloeden (Monks et al. 2016). De mechanismen die de verjonging van bomen reguleren zijn echter nog niet zo goed gekend. Er zijn studies die suggereren dat de reproductie van bomen in de toekomst kan toenemen, terwijl anderen veronderstellen dat de reproductie kan afnemen als reactie op het veranderend klimaat (Allen et al. 2014; Redmond et al. 2012). In Vlaanderen stellen we vast dat de mastjaren bij eik en beuk elkaar steeds korter opvolgen. Vroeger werd aangenomen dat beuken slechts om de 5 à 12 jaar een zogenaamd zaadjaar kenden. In een zaadjaar of mastjaar zijn er opvallend veel bomen met zaadproductie, terwijl in de tussenjaren weinig of geen zaden worden gevormd. Onder invloed van de klimaatopwarming blijkt de beuk steeds vaker zaad te produceren. Om de 2 à 3 jaar een mastjaar is niet meer uitzonderlijk (Sioen & Verstraeten, 2023). Om meer vat te krijgen op deze onzekerheden, zijn experimenten waarin planten worden blootgesteld aan de voorspelde groeicondities essentieel. In dit kader onderzochten wij de effecten van een ernstige droogteperiode op de reproductieve eigenschappen van de struiksoort sporkehout (synoniemen: spork, vuilboom; wetenschappelijke naam: Frangula alnus, oudere wetenschappelijke naam: Rhamnus frangula).
Materiaal en methoden
Sporken hebben het grote voordeel dat de planten al op jonge leeftijd bessen produceren, zelfs wanneer ze in potten worden opgekweekt. Zo kunnen we een eenvoudige droogteproef uitvoeren op jonge, reproductieve potplanten. Bij boomsoorten zoals beuk of eik is het veel moeilijker een dergelijke manipulatieve proef uit te voeren. Hoe groter de boom, hoe groter ook de benodigde pot. Tegen dat deze bomen zaden produceren zou je al een heftruck nodig hebben om de pot te verplaatsen… Hoewel spork als struiksoort geen economische waarde heeft, er wordt geen hout van geoogst voor zagerijen of industriële verwerking, vervult het wel een ecologische rol in onze bossen.
We onderzochten sporken afkomstig uit natuurlijke populaties in Italië, België en Zweden (telkens 1 populatie). De populaties bevonden zich allen in laagland. De zaden werden lokaal geoogst en in onze kwekerij te Geraardsbergen opgekweekt tot potplanten. Van deze zaailingen werden stekken gesneden en opgekweekt, zodat we voor elke zaailing ook klonen bekwamen. Zo konden we genetisch identieke individuen met elkaar vergelijken bij droogte en in controle. Onze proefopzet was een zogenaamde “common garden”. Hierbij worden alle planten onder dezelfde condities opgekweekt, en worden de verschillende herkomsten individueel gemengd, zodanig dat verschillen tussen de herkomsten niet te wijten kunnen zijn aan verschillen in groeiomstandigheden – want die zijn voor alle planten dezelfde – maar wel aan genetische verschillen. Zowel voor, tijdens als na de droogteproef waren de drie herkomsten steeds individueel en at random gemengd. Voor de volledige beschrijving van materiaal en methode verwijzen we naar de wetenschappelijke artikels (Vander Mijnsbrugge et al. 2020, Vander Mijnsbrugge et al. 2022).


Figuur 1: links: bloei bij spork, rechts: besvorming bij spork
Initieel werden de sporken buiten, op een containerveld opgekweekt. In de zomer van 2018 plaatsten we de jonge potplanten in een serre, te Geraardsbergen, en onderwierpen een gedeelte van de planten aan een ernstige droogteperiode door de planten helemaal geen water te geven gedurende 28 dagen (Vander Mijnsbrugge et al. 2020). Een ander deel van de planten, de controles, kreeg wel water (Fig. 2). Het verdrogen van de bladeren, waarbij het bladweefsel eerst soepelheid verliest en naar beneden begint te hangen, vervolgens verkleurt en opkrult om ten slotte helemaal te verwelken, werd nauwkeurig opgevolgd (Fig. 3). Sommige planten overleefden de droogteperiode niet. Na het droogte-experiment werden de controle en droogteplanten gemengd en terug buiten, op het containerveld geplaatst (Fig. 4).

Figuur 2: opstelling van de droogteproef met controle (a) en uitgedroogde planten (b)





Figuur 3: verschillende stadia van uitdroging bij spork

Figuur 4: na de droogtestress: sporken in pot op het containerveld
Resultaten
De eerste resultaten van deze proef waren contra-intuïtief. Van de Italiaanse herkomst verwachtten we dat deze beter aangepast was aan hogere temperaturen in het groeiseizoen en dat deze dan ook droogteperiodes beter zou kunnen doorstaan. Deze herkomst vertoonde evenwel als eerste droogtesymptomen tijdens de droogteperiode. Uiteindelijk stierven meer planten van de Italiaanse herkomst af. De reden was vermoedelijk het feit dat de Italiaanse herkomst gemiddeld gezien grotere bladeren had en deze bladeren ook een groter aantal huidmondjes telden per oppervlakte-eenheid. Huidmondjes zorgen voor gasuitwisseling, het zijn heel kleine openingen in het blad. Door de huidmondjes haalt de plant overdag koolstofdioxide (CO2) binnen uit de lucht, legt de koolstof (C) vast via fotosynthese, en laat zuurstof (O2) weer naar buiten. Maar door deze openingen verliest de plant ook vocht. De plant kan de huidmondjes sluiten, maar als deze te laat in actie schiet met sluiten loopt het mis. Planten laten normaal huidmondjes zo lang mogelijk open om de fotosynthese zo weinig mogelijk te hinderen. Dit alles zorgde er vermoedelijk voor dat de planten van de Italiaanse herkomst sneller vocht verloren.
Twee jaar na de behandeling vertoonden de droogte-planten nog steeds een kleinere hoogtegroei dan de controleplanten (Fig. 5). Het is gemakkelijk te begrijpen dat droogte de groei van houtige planten vertraagt. Alleen al het sluiten van de huidmondjes onder drogere condities hindert reeds de fotosynthese. Het is opvallend dat deze groeivertraging zich ook (soms) doorzet in de jaren na de droogte.
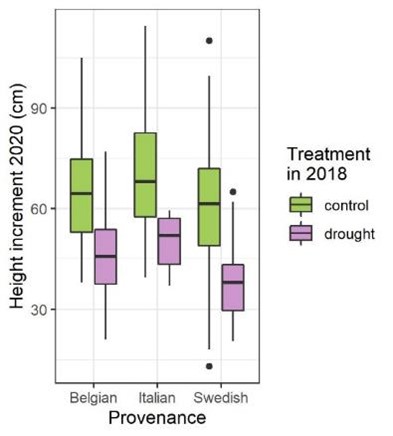
Figuur 5: aanwas in hoogte voor de drie verschillende herkomsten (lokaal, Italiaans en Zweeds) twee jaar na de droogtebehandeling voor zowel controle als gestresseerde planten
De hamvraag hebben we hier echter nog niet beantwoord: wat er gebeurde met de reproductieve kenmerken van de sporken. We hebben de rijpe bessen die op de planten gevormd werden geteld op regelmatige tijdstippen in het groeiseizoen, zowel één jaar als twee jaar na de droogte (Vander Mijnsbrugge et al. 2022). Omdat spork gedurende de volledige zomer continu bessen aanmaakt, van eind juni tot in september, doch met een piek midden juli, werden telkens de rijpe bessen op alle planten geteld en tevens geplukt zodat in een volgende telronde de eerder getelde bessen niet opnieuw werden meegerekend. Twee jaar na de droogtebehandeling hebben we de getelde en geplukte bessen ook bijgehouden, de zaden uit de bessen gehaald, gedroogd, gewogen en doen kiemen (Fig. 6).


Figuur 6: links: heel wat manueel werk om bessen te plukken op de afzonderlijke planten, en dit op verschillende tijdstippen in het groeiseizoen; rechts: vele partijtjes zaden kiemen
Op basis van het aantal geoogste bessen doorheen het jaar stelden we vast dat de planten die droogte ondergaan hadden één en twee jaar na de behandeling meer bessen produceerden in vergelijking met de controleplanten en dit gedurende het hele groeiseizoen (figuur). Midden juli produceerde een droogte-behandelde moederplant gemiddeld een 15 tot 20% meer bessen dan de controle. Naast de hogere besproductie voor de droogte-behandelde planten, vertoonde de Zweedse herkomst iets vroeger een piek in besproductie dan planten uit België en Italië (figuur). De bladontwikkeling vanuit de overwinterende knoppen van de Zweedse herkomst in het voorjaar gebeurde ook iets vroeger dan bij de Vlaamse herkomst, maar de Zweedse herkomst vertoonde vooral op het eind van het groeiseizoen een flink vroegere bladverkleuring en bladval. De Italiaanse herkomst liep ook wel wat vroeger uit in het voorjaar dan de Vlaamse herkomst, maar vertoonde geen vervroegde bladverkleuring in het najaar. Ook interessant is dat de Italiaanse sporken zowel bij de controle als bij de behandelde planten minder bessen produceerden dan de Vlaamse herkomst (Fig. 7).
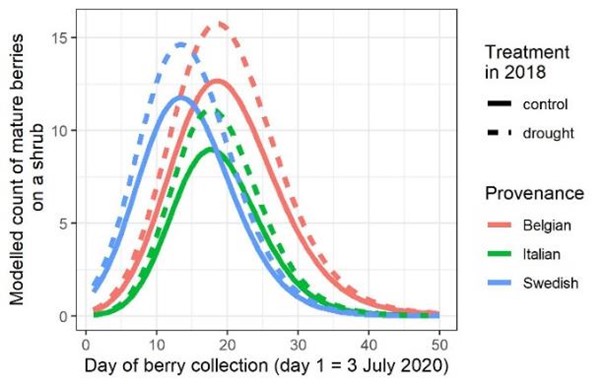
Figuur 7: gemodelleerd aantal bessen geproduceerd op de verschillende herkomsten (lokaal, Italiaans en Zweeds) twee jaar na de droogtebehandeling voor zowel de controle als de gestresseerd planten, gedurende de zomer (juli en augustus)
Het gemiddelde gewicht van de zaden uit de bessen, geplukt twee jaar na de droogtebehandeling, en hun kiemingspercentage waren hoger voor de droogte-behandelde planten dan voor de controleplanten. Gemiddeld woog een zaadje rond de 18 microgram. Een kleine 1 microgram verschil zat er tussen controle en droogte-moederplanten (Tabel 1). Een klein doch significant verschil. Mogelijk reflecteert dit resultaat een algemene strategie in het plantenrijk waarbij tolerantie tegen droogte en zwaardere zaden dikwijls samen voorkomen (Stahl et al. 2013). Een zwaarder zaad laat een vlugge kiemwortelvorming toe die sneller doorheen droge oppervlakkige bodemlagen kunnen heen groeien (Leishman etal. 1994). Het gewicht van de zaden vertoonde, net zoals het aantal bessen op een plant, een piek voor de oogstdag in de tweede helft van juli. De laatste oogstdag, een maand later, liet veel lagere zaadgewichten optekenen.
Tabel 1: gemiddeld gewicht van een zaadje en gemiddeld kiempercentage berekend per oogstdag voor zowel controle als gestresseerde moederplanten, twee jaar na de droogtebehandeling
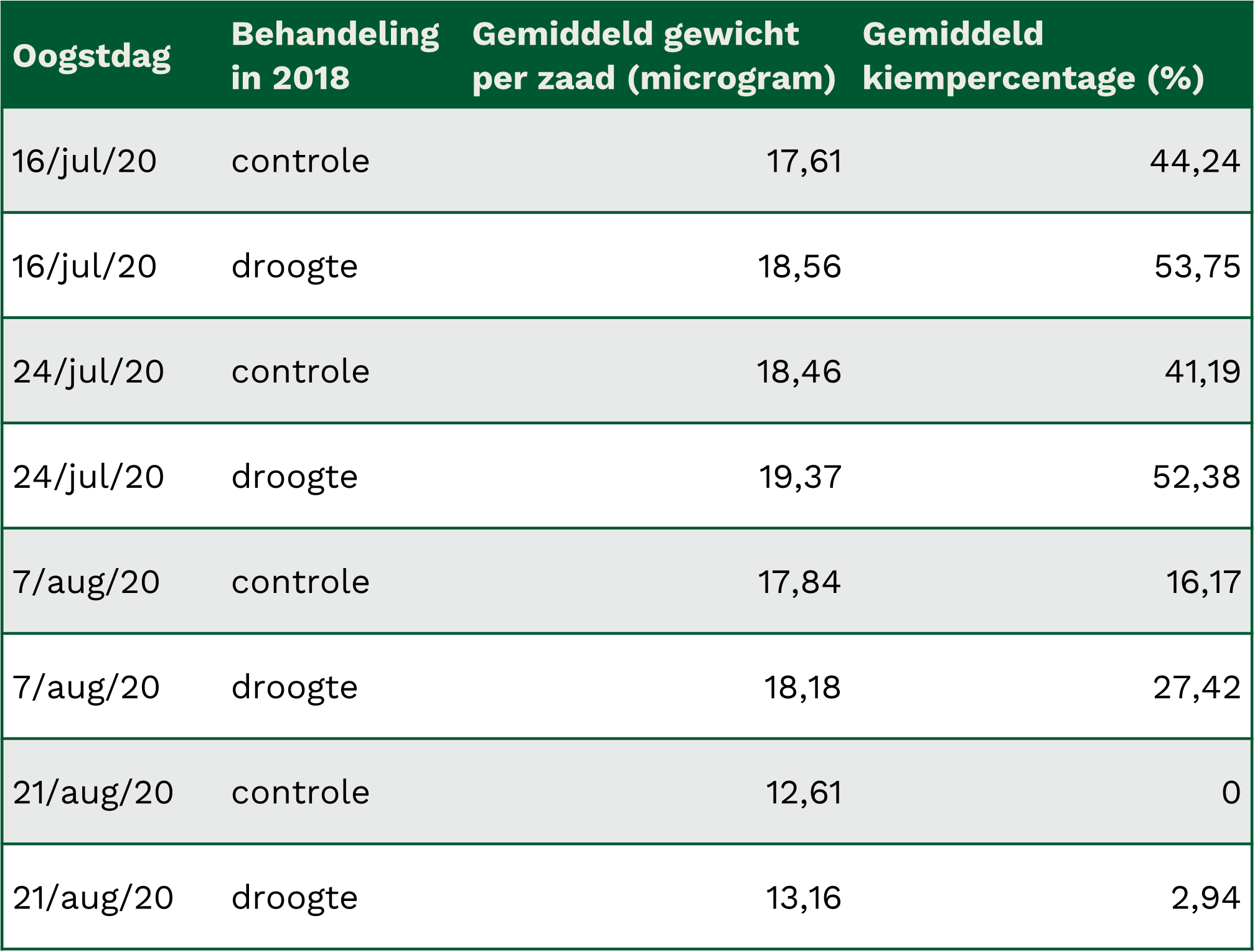
Het gemiddeld kiempercentage van de zaden lag in juli tussen de 40% en 55%, en zakte drastisch naarmate het groeiseizoen vorderde, met nagenoeg geen kieming meer voor de laatst geoogste zaden (Tabel 1). Zaden van droogte-behandelde moederplanten vertoonden een tot 10 % hoger kiempercentage.
Het hogere kiemsucces van de zaden geplukt op de droogte-behandelde planten twee jaar na de droogteperiode is een interessant fenomeen. Het wordt een transgenerationeel effect genoemd, omdat gevolgen van iets wat de moederplant heeft ondergaan nog waarneembaar zijn in de nakomelingen (Herman et al. 2011). De moederplant “onthoudt” wat er gebeurd is en geeft het door aan de nakomelingen. Het iets hogere kiempercentage wijst erop dat de moederplant wat meer investeert in de zaden opdat de hieruit voortkomende zaailingen over betere overlevingskansen kunnen beschikken, wat zinvol is in een stressvollere omgeving. Een hoger kiempercentage na droogte is een gekend fenomeen bij kruidachtige planten (Mojzes et al. 2021). Zowel voor zaadgewicht als voor kiempercentage zagen we geen opvallende verschillen tussen de drie herkomsten die we onderzochten.
Discussie
Samen met de mindere groei wijzen deze resultaten erop dat planten die droogte ondergaan hebben, tot twee jaar na de droogte meer energie steken in reproductie dan de controleplanten en minder in groei dan de controles. Een analoog resultaat werd vastgesteld in een droogteproef op steeneik en aardbeiboom in een mediterrane omgeving waar na 18 jaar verminderen van regen door afscherming van de boomkronen een verhoogde reproductie werd genoteerd (Bogdziewicz et al. 2020). Het is aannemelijk dat bij condities die effectief stress veroorzaken een trade-off tussen groei en reproductie optreedt. In een jaarringanalyse op beuk stelde men vast dat de grootste groeireductie zich voordeed in jaren met zowel sterke zaadproductie gecombineerd met een stevige droogteperiode in de zomer (Hacket-Pain et al. 2017). Door verhoudingsgewijs meer te investeren in reproductie tracht de boom de kans te verhogen op succesvolle nakomelingen, waarbij de boom tegelijkertijd wel de kans op zelf afsterven kan verhogen gezien mindere hulpbronnen naar groei kan samengaan met beperktere beschermingsmechanismen tegen stress (Lauder et al. 2019).
We observeerden ook het moment dat de zaailingen kiemden in het voorjaar van 2021. Daaruit bleek dat zaden afkomstig van bessen geoogst later op het groeiseizoen een beetje vroeger kiemden. Zaden geoogst op 21 augustus kiemden ongeveer 4 dagen vroeger dan zaden geoogst op 16 juli. En als we naar de herkomst van de moederplant keken bleek dat zaden van zowel de Italiaanse als de Zweedse herkomst ongeveer 4 à 5 dagen vroeger kiemden dan de Vlaamse herkomst. Dit patroon vonden we ook terug bij het uitlopen van de knoppen op de moederplanten in het voorjaar: de Italiaanse en Zweedse herkomst lopen beiden systematisch vroeger uit dan de Vlaamse herkomst. Het lijkt er dus op dat er een gemeenschappelijke genetische basis bestaat voor beide fenologische kenmerken: moment van kieming en moment van knopuitloop in het voorjaar (Rohde et al. 2007). Wel stelden we vast dat knopuitloop in het voorjaar sterker genetisch bepaald is dan het tijdstip van kieming. Dit lijkt ook niet zo onlogisch. Kieming is een once-in-a-lifetime gebeurtenis. Hier mag niets mislopen. Het proces moet zich perfect op de lokale microcondities kunnen afstemmen en dus minder genetisch gedetermineerd zijn. Die nood is minder sterk bij knopuitloop. Zolang de plant niet afsterft kunnen nieuwe knoppen ontwikkelen en uitlopen bijvoorbeeld na schade door een late voorjaarsvorst.
Het aantal rijpe bessen, het gewicht van de zaden in deze bessen en het kiempercentage van de zaden daalden na het piekmoment halverwege juli. Dit roept de vraag op waarom een plant investeert in besproductie na deze piekperiode, wanneer de kans op gevestigde zaailingen vermindert. Mogelijk is dit een strategie van risicospreiding. In het geval van slechte omstandigheden voor besproductie tijdens de piekperiode en betere erna, is er toch nog mogelijkheid om voor levensvatbare zaailingen te zorgen, ook al zal het aantal en de kiemkracht lager zijn.
Onze resultaten suggereren dat geassisteerde migratie, waarbij we herkomsten van bomen en struiken afkomstig uit meer zuidelijk en/of oostelijk gelegen regio’s in Europa bij ons aanplanten als een anticipatie op de klimaatverandering, niet altijd de verwachte patronen volgt. Laten we waakzaam blijven.
Met dank aan…
Heel wat mensen hebben bijgedragen tot dit onderzoek, onmogelijk om iedereen te vernoemen. Hieruit wel een greep: Hanne zorgde voor het initiële zaad, Stefaan heeft de zaden gestratificeerd, laten kiemen en verder opgekweekt en verzorgd; Lise, Nele en Amy hebben de droogteproef uitgevoerd en opgevolgd; Laura, Marie en ook Yorrick hebben vele bessen geteld en geplukt; Stefaan, Marc en Yorrick hebben vele bessen geschoond, gewogen en gestratificeerd; Denis en Stijn hebben vele zaailingen geteld.
Gelieve als volgt te citeren:
Kristine Vander Mijnsbrugge (2024) Invloed van droogte op reproductie bij spork. Bosrevue 112a
ISSN 2565-6953 – Bosrevue 112a
-
Referenties
Patacca, M.; Lindner, M.; Lucas-Borja, M.E.; Cordonnier, T.; Fidej, G.; Gardiner, B.; Hauf, Y.; Jasinevičius, G.; Labonne, S.; Linkevičius, E.; et al. Significant increase in natural disturbance impacts on European forests since 1950. Global Change Biology n/a, doi:https://doi.org/10.1111/gcb.16531.
Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management 2010, 259, 660-684, doi:https://doi.org/10.1016/j.foreco.2009.09.001.
Monks, A.; Monks, J.M.; Tanentzap, A.J. Resource limitation underlying multiple masting models makes mast seeding sensitive to future climate change. The New phytologist 2016, 210, 419-430, doi:10.1111/nph.13817.
Redmond, M.D.; Forcella, F.; Barger, N.N. Declines in pinyon pine cone production associated with regional warming. Ecosphere 2012, 3, 1-14.
Sioen, G.; Verstraeten, A. Geen vruchtbaar jaar voor zomereik, wel heel wat beukennootjes. INBO nieuwsbrief, november 2023.
Vander Mijnsbrugge, K.; De Clerck, L.; Van der Schueren, N.; Moreels, S.; Lauwers, A.; Steppe, K.; De Ligne, L.; Campioli, M.; Van den Bulcke, J. Counter-Intuitive Response to Water Limitation in a Southern European Provenance of Frangula alnus Mill. in a Common Garden Experiment. Forests 2020, 11, 1186.
Vander Mijnsbrugge, K.; Schouppe, M.; Moreels, S.; Aguas Guerreiro, Y.; Decorte, L.; Stessens, M. Severe Drought Still Affects Reproductive Traits Two Years Later in a Common Garden Experiment of Frangula alnus. Forests 2023, 14, 857.
Allen, R.B.; Hurst, J.M.; Portier, J.; Richardson, S.J. Elevation-dependent responses of tree mast seeding to climate change over 45 years. Ecol Evol 2014, 4, 3525-3537, doi:10.1002/ece3.1210.
Herman, J.; Sultan, S. Adaptive Transgenerational Plasticity in Plants: Case Studies, Mechanisms, and Implications for Natural Populations. Frontiers in Plant Science 2011, 2, doi:10.3389/fpls.2011.00102.
Bogdziewicz, M.; Fernández-Martínez, M.; Espelta, J.M.; Ogaya, R.; Penuelas, J. Is forest fecundity resistant to drought? Results from an 18-yr rainfall-reduction experiment. The New phytologist 2020, 227, 1073-1080, doi:10.1111/nph.16597.
Caignard, T.; Kremer, A.; Firmat, C.; Nicolas, M.; Venner, S.; Delzon, S. Increasing spring temperatures favor oak seed production in temperate areas. Scientific Reports 2017, 7, 8555, doi:10.1038/s41598-017-09172-7.
Hacket-Pain, A.J.; Lageard, J.G.A.; Thomas, P.A. Drought and reproductive effort interact to control growth of a temperate broadleaved tree species (Fagus sylvatica). Tree Physiology 2017, 37, 744-754, doi:10.1093/treephys/tpx025.
Lauder, J.D.; Moran, E.V.; Hart, S.C. Fight or flight? Potential tradeoffs between drought defense and reproduction in conifers. Tree Physiology 2019, 39, 1071-1085, doi:10.1093/treephys/tpz031.
Stahl, U.; Kattge, J.; Reu, B.; Voigt, W.; Ogle, K.; Dickie, J.; Wirth, C. Whole-plant trait spectra of North American woody plant species reflect fundamental ecological strategies. Ecosphere 2013, 4, art128, doi:https://doi.org/10.1890/ES13-00143.1.
Leishman, M.R.; Westoby, M. The Role of Seed Size in Seedling Establishment in Dry Soil Conditions — Experimental Evidence from Semi-Arid Species. Journal of Ecology 1994, 82, 249-258, doi:10.2307/2261293.
Mojzes, A.; Kalapos, T.; Kröel-Dulay, G. Drought in maternal environment boosts offspring performance in a subordinate annual grass. Environmental and Experimental Botany 2021, 187, 104472, doi:https://doi.org/10.1016/j.envexpbot.2021.104472.
Rohde, A.; Bhalerao, R. Plant dormancy in the perennial context. Trends in plant science 2007, 12, 217-223, doi:10.1016/j.tplants.2007.03.012.



