


Bosrevue

Heel wat beuken, vooral in het open landschap en langs bosranden, hebben het lastig met de hitte en de droogte van de voorbije jaren. Door dit opvallende beeld verschijnen regelmatig doemberichten: de beuk zou vooraan in het rijtje staan van soorten die uit onze streken dreigen te verdwijnen door klimaatverandering.
Bovendien heeft de beuk geen al te goede reputatie op het vlak van biodiversiteit: beuk zou door zijn hoge schaduwdruk en slecht afbreekbaar strooisel de biodiversiteit van onze bossen bedreigen. Sommige beheerders kappen daarom gaten in beukenbossen om andere boomsoorten, zoals wintereiken, in de plaats aan te planten. We plaatsen bij deze aanpak een aantal kanttekeningen op basis van de wetenschappelijke literatuur over beuk en beukenbossen.
De ongekroonde koning van ons bos
Beuk is een Europese boomsoort die voorkomt van het noorden van Spanje en de Balkan tot in Zweden en Noorwegen. Vlaanderen ligt pal in het midden van deze zuid-noord verspreiding en in vergelijking met andere regio’s halen beuken hier bijzonder grote dimensies (Vandekerkhove et al., 2018a). Toch is er in onze regio (met uitzondering van het Zoniënwoud) geen diep gewortelde traditie in het beheer van beukenbossen zoals in Centraal-Europa. Onze bossen werden immers traditioneel als hakhout of middelhout beheerd en beuken laten zich in onze streken moeilijk in hakhout zetten. Bovendien laat een bovenetage van beuk weinig licht door, zodat er onder de beuken ook weinig hakhout kan groeien. Deze boomsoort werd dan ook maar met mondjesmaat getolereerd in onze bossen. In de 19de eeuw werd het hak- en middelhoutbeheer geleidelijk aan verlaten. Op de minst productieve, zandige plaatsen werd het omgevormd naar hooghout van beuk of naaldhout (Vandekerkhove et al., 2016). In bossen die voorheen als middelhout werden beheerd, neemt beuk spontaan toe nadat dit middelhoutbeheer gestaakt is. Dat is niet alleen bij ons het geval, maar in vele Europese bossen (Tolksdorf et al., 2015; Dupin et al., 2017; Máliš et al. 2020). In onze regio is het Zoniënwoud altijd een buitenbeentje geweest: beuk was er al minstens sinds het jaar 600 dominant aanwezig (Deforce et al., 2018) en het bos wordt al sinds de middeleeuwen als een spontaan verjongend, opgaand beukenbos beheerd (Vandekerkhove et al., 2018b). Dankzij de hoge natuurlijkheid en de bijzondere biodiversiteit, zijn spontaan ontwikkelende reservaten in het Zoniënwoud een referentie voor natuurlijke beukenbossen in onze streken. De bosreservaten van het Zoniënwoud kregen daarom erkenning als UNESCO werelderfgoed.
De beuk en klimaatverandering
Door klimaatverandering wordt het gemiddeld warmer en neemt de lengte van het groeiseizoen toe. Wellicht zal er jaarlijks meer neerslag vallen, maar vooral dan in de winter, terwijl er in het groeiseizoen langere periodes van droogte afwisselen met periodes van intense regenval (Mira, 2015; https://www.meteo.be/nl/info/nieuwsoverzicht/wat-houdt-de-langdurige-droogte-in-stand). Maar over heel wat andere effecten van klimaatverandering is nog veel onzekerheid. Zo verwacht men meer instabiliteit met afwisselend warme periodes en vorst in het voorjaar. Daardoor kan er frequenter vorstschade in het voorjaar optreden (Zohner et al, 2017). Eén ding is zeker: wat op ons afkomt is een ‘nieuw’ klimaatbeeld, niet het klimaat van ergens anders dat gewoon wat opschuift (Boussemaere & Vicca, 2020).
We kunnen moeilijk voorspellen welk klimaat we over 100 jaar gaan kennen en nog moeilijker inschatten wat de plasticiteit en weerbaarheid is van de verschillende boomsoorten in dat nieuwe klimaat. Ook weten we niet goed in welke mate gesloten bossen een bufferend effect hebben en hoe specifiek klimaatverandering inwerkt op verschillende groeiplaatsen.
Het hoeft dan ook niet te verbazen dat prognoses over de toekomstige geschiktheid van boomsoorten en areaalveranderingen door klimaatverandering, geen eenduidig beeld geven voor heel wat soorten. Dat is zeker ook het geval voor beuk: sommige studies voorspellen een aanzienlijke achteruitgang in het natuurlijke areaal dat geschikt is voor beuk (Meier et al., 2012; Maiorano et al., 2013; Thurm et al. 2018), terwijl andere beuk zien als één van de grote winnaars, die zijn areaal fors kan uitbreiden (Falk & Hempelmann, 2013; Dyderski et al., 2018). We illustreren dit met de resultaten van twee recente studies (Dyderski et al., 2018; Thurm et al., 2018), die dezelfde klimaatscenario’s hanteren maar toch tot sterk uiteenlopende prognoses komen. Volgens Thurm et al. (2018) wordt Vlaanderen tegen 2061-2080 nauwelijks nog geschikt voor beuk bij een temperatuurstijging van 2.9°C (klimaatscenario RCP 4.5), maar kan de soort zich handhaven in de koelere Ardennen. Bij een temperatuurstijging van 4.5°C (klimaatscenario RCP 8.5) verschuift het areaal van beuk volgens deze studie helemaal naar Alpiene regio’s, het noorden van de Britse eilanden en Scandinavië en wordt Vlaanderen zo goed als ongeschikt (figuur 1). De studie van Dyderski et al. (2018) geeft enkel verschuivingen in het areaal weer, maar niet de geschiktheid. Deze studie voorspelt eveneens dat beuk zich vestigt in montane en noordelijke gebieden, terwijl de soort verdwijnt in zuidelijke en oostelijke delen van zijn areaal. Vlaanderen blijft volgens Dyderski et al. (2018) ook volgens het meest pessimistische scenario (RCP 8.5) binnen het areaal van beuk liggen (figuur 2).
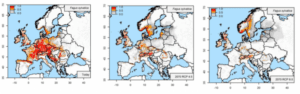
Figuur 1: Geschiktheid (0 = ongeschikt; 1 = zeer geschikt) in het huidige klimaat (links) en toekomstige geschiktheid voor Beuk bij klimaatscenario’s RCP 4.5 (midden) en RCP 8.5 (rechts) volgens Thurm et al. (2018).

Figuur 2: Huidige (a) en toekomstige arealen van beuk bij een optimistisch (b), gemiddeld (c) en pessimistisch (d) scenario volgens Dyderski et al. (2018). Rood is de afname van het toekomstige areaal, blauw de toename. Groen is de overlap tussen het actuele voorkomen en het toekomstige areaal.
De hierboven besproken scenario’s houden geen rekening met veranderingen in het land- en bosbeheer, die kunnen interfereren met de effecten van klimaatverandering (Bodin et al., 2013; Vayreda et al., 2016). Zo neemt de sluitingsgraad en de houtvoorraad in onze bossen al enkele decennia toe en toonden Von Arx et al. (2013) en Zellweger et al. (2020) aan dat het bufferende microklimaat binnen het bos versterkt naarmate de biomassa en de dichtheid van het kronendak toeneemt. De voorspellende modellen die hierboven werden besproken, gebruiken de relaties tussen de boomsoort en zijn leefomgeving om in de toekomst te kijken. Deze relaties zijn echter een veranderend samenspel van diverse aspecten van het klimaat met het bosbeheer, de bodemeigenschappen e.a. Verschillende aannames over het toekomstige samenspel tussen deze factoren, verklaren wellicht de uiteenlopende prognoses voor beuk in onze regio. Aangezien deze modellen geen uniforme uitkomst geven en er nog zo veel onzekerheden zijn, is het niet aangewezen om er nu al verregaande consequenties aan te verbinden voor het beheer van onze beukenbossen (zie bv. Daise et al., 2009).
Vitaliteit en stabiliteit van beukenbossen
Jaarringanalyses wijzen uit dat de groei van beuken in Zoniënwoud en Meerdaalwoud toenam tussen 1875 en 1960 en daarna geleidelijk aan terug verminderde (Aertsen et al., 2014). Een gelijkaardige trend wordt ook in Centraal-Europa waargenomen (Härdtle et al., 2013). De toename van de groei in de eerste periode wordt vooral toegeschreven aan veranderingen in het bosbeheer en aan een hogere beschikbaarheid van stikstof, onder meer door atmosferische depositie. De groeivertraging na 1990 kan toegeschreven worden aan toegenomen droogtestress en hittegolven in het voorafgaande jaar (Härdtle et al., 2013; Latte et al., 2015). Droogte heeft tot gevolg dat de groei van beuk aanzienlijk vertraagt, maar dit kan een fysiologische aanpassing zijn die niet per definitie samengaat met vitaliteitsproblemen (Gessler et al., 2020). Ondanks de vertraging was de groei in het Meerdaalwoud in 2008 nog altijd 15% hoger dan in 1900 en in het Zoniënwoud bedroeg de bonus sinds 1900 nog 8%. Op beide zeer productieve groeiplaatsen is de groeivertraging kleiner dan elders in Europa, op minder geschikte groeiplaatsen (Aertsen et al., 2014).
Terwijl heel wat beuken in het open landschap zichtbaar schade hebben opgelopen door de droogte en de hitte van de voorbije jaren, hebben beuken in bossen bij ons voorlopig blijkbaar niet meer problemen dan andere boomsoorten (figuur 3). Dat blijkt ook uit de jaarlijkse steekproef van de bosvitaliteit in Vlaanderen. Beuken vertoonden in de periode 1995-2019 minder bladverlies in de zomer dan andere veel voorkomende boomsoorten, zoals zomereik. Wel is er voor beuk in deze periode een licht stijgende trend in het bladverlies in de zomer waarneembaar, wat niet het geval is voor zomereik (Sioen et al., 2020). Tijdens mastjaren, dit zijn jaren waarin de bomen veel meer zaad vormen dan in gewone jaren, is de bladbezetting wel lager. Mastjaren komen bij beuk nu meer voor dan vroeger en ook het uitzonderlijk droge jaar 2018 was een mastjaar. In 2019 verbeterde echter de kroonconditie van beuk, dit in tegenstelling tot die van zomereik (Sioen et al., 2020).


Figuur 3:Terwijl beuken in het open landschap er vaak slecht aan toe zijn na drie droge en hete zomers (foto boven), houden beuken in gesloten bossen goed stand (foto onder). Beide foto’s zijn op enkele 100den meter afstand van elkaar genomen (Kruiskerke, Ruiselede; Foto’s: Kris Vandekerkhove, oktober 2020)
Mortaliteitsgegevens van natuurlijke, onbeheerde beukenbossen in Zweden, Oekraïne, Zwitserland en Duitsland geven nog geen sluitend bewijs van verhoogde sterfte bij volwassen beuken in de voorbije jaren. Onderlinge competitie, windval en aantastingen zijn in deze bossen tot op heden een betere verklaring voor sterfte dan droogte. Lokaal stellen zich wel problemen, vooral op een ondiepe bodem. Op dergelijke groeiplaats in het nationaal park Hainich (Thüringen, centraal in Duitsland) stierf na langdurige droogtes in 2018 en 2019, 30% van de oude beuken af (Leuschner, 2020).
Ondiepe bodems kunnen tijdelijk zeer nat zijn, maar ook sterk uitdrogen. Op deze bodems vormt beuk enkel oppervlakkige wortels, waardoor de soort er niet alleen gevoelig voor uitdroging, maar ook voor omwaaien (Nabuurs & Hommel, 2007). Problemen gerelateerd aan oppervlakkige beworteling komen veel minder voor op goed doorlatende zand- of leembodems. Daar vormt beuk een dubbel wortelstelsel, met oppervlakkige wortels waarmee hij de bovenste bodemlagen inneemt, maar daarnaast ook met metersdiepe wortels, die voldoende stabiliteit bieden en die voedingsstoffen en water uit de diepere bodem naar boven halen (Langohr, 2009; Meier et al. 2018).
In vele Vlaamse bossen, zowel op zand- als op leemgronden, zien we regelmatig jonge, natuurlijk gevestigde beuken die ook verder doorgroeien (Fig. 4). In ons grootste beukenbos, het Zoniënwoud, zien we sinds enkele decennia veel meer gevestigde en doorgroeiende verjonging van beuken dan voorheen, zonder duidelijk aanwijsbare reden (Everts et al., 2020). Deze succesvolle verjonging en het feit dat de jonge beuken weinig hinder schijnen te ondervinden van de droge periodes van de afgelopen jaren, kunnen er op wijzen dat de actuele omstandigheden in vele van onze bossen nog steeds geschikt zijn voor beuk.

Figuur 4: Jonge beuken die zich spontaan hebben gevestigd in een naaldhoutbestand op zandgrond, hebben de voorbije droogte en hitte goed doorstaan en groeien door naar de nevenetage (Drongengoed, oktober 2020; foto Luc De Keersmaeker).
Samen sterk
De droogte en hitte van de voorbije zomers hebben veel grotere effecten gehad op beuken in het open landschap en langs bosranden dan in gesloten bossen (figuur 3). Deze opvallende verschillen zijn minstens gedeeltelijk te verklaren door de kenmerken van beuk. Beuk is een uitgesproken climaxboomsoort van koele en donkere bossen, die minder goed bestand is tegen een combinatie van hoge temperatuur, hoge lichtintensiteit en lage luchtvochtigheid dan lichtbehoevende soorten. Anders dan bij eik en es, biedt bij beuk niet zozeer de bouw en samenstelling van de bladeren, maar wel de kroonarchitectuur bescherming tegen extreem hoge instraling (Kutsch et al. 2009). Beuken hebben een zeer grote totale bladoppervlakte en de kronen groeien onder of in elkaar. Hierdoor is een aanzienlijk deel van de bladeren en de schors beschaduwd door andere bladeren, van dezelfde boom of van een naburige boom. Dit beschermt de schors en het fotosynthesapparaat van beuk, dat gevoeliger blijkt te zijn voor hoge temperaturen dan dat van een pionier als berk (Peltzer, 2001). Beuken zijn op bestandsniveau relatief zuinig met water: ze verbruiken een vergelijkbare hoeveelheid als zomereik, wintereik en fijnspar, maar minder dan uitgesproken pionierboomsoorten zoals ruwe berk en grove den (Leuschner & Ellenberg, 2017). Jonge beuken zijn gevoelig voor uitdroging maar ze kunnen, meer dan vele andere inheemse boomsoorten zoals zomereik, vocht opnemen via de bladeren om een vochtdeficit te voorkomen (Schreel et al., 2019). Dit is wellicht een aanpassing aan een getemperd vochtig microklimaat, zoals dat in gesloten bossen heerst. Daarnaast is ook een aanzienlijke herverdeling van water vastgesteld tussen naburige beukjes, die al of niet via mycorrhiza wortelcontact maken (Hafner et al., 2020). De associatie met ectomycorrhiza stelt beuken in staat op een efficiënte wijze voedingsstoffen te ontsluiten en het belang van een aangepaste mycorrhizagemeenschap lijkt toe te nemen bij droogtestress (Shi et al., 2002; Pena & Polle, 2014; Leberecht et al., 2015; Nickel et al., 2018). Verschillende bomen, vooral van dezelfde of een verwante soort, kunnen via mycorrhiza onderling ook aanzienlijke hoeveelheden koolstofverbindingen uitwisselen (Klein et al, 2016; Rog et al., 2020). De associatie van mycorrhiza met zaailingen wordt door het bosbeheer beïnvloed: in bestandsopeningen is de diversiteit van mycorrhiza veel lager dan in gesloten beukenbos (Grebenc et al., 2009).
Dit alles illustreert dat beuken in bosverband bijzonder performant zijn en hun eigen optimale groeiomstandigheden creëren en in stand houden, waardoor ze in natuurlijke bossen kunnen domineren. Wanneer deze voordelen ontbreken (dreefbomen in het open landschap) of door verstoringen wegvallen (overstaanders of randbomen bij kapvlakten), dan blijken beuken juist heel kwetsbaar te zijn.
De biodiversiteit van beukenbossen
Beukenbossen worden geassocieerd met een lage biodiversiteit: het zijn vaak donkere bossen met weinig ondergroei, en er zijn op het eerste zicht heel weinig planten of dieren te zien. Toch kan de soortenrijkdom van beukenbossen juist zeer groot zijn, maar die is vooral terug te vinden in onopvallende soortengroepen zoals zwammen, mossen en ongewervelden die leven in en op dood hout. Daar zijn ook heel wat soorten bij die specifiek gebonden zijn aan beuk (Assmann et al., 2007; Ódor et al., 2005; Meyer & Schmidt, 2008; Brunet & Isacsson, 2009; Fritz, 2009; Fritz et al., 2009; Fritz & Heilmann-Clausen, 2010; Heilmann-Clausen & Christensen, 2003; Christensen et al., 2005; Moning & Müller, 2009; Lachat et al., 2012; Speight, 1989).
Onderzoek op doodhoutkevers (Müller et al., 2012) toonde aan dat ruim 70% van alle gekende saproxyle keversoorten in Europa reeds in door beuken gedomineerde bossen is aangetroffen. Verder blijkt dat vooral oude beukenbestanden en –bomen zeer soortenrijk zijn. Heel wat zeldzame en veeleisende soorten kevers, mossen, korstmossen en zwammen komen hoofdzakelijk of uitsluitend op deze oude bomen voor (Fig. 5).

Figuur 5: In structuurrijke, natuurlijk ontwikkelende beukenbossen hebben monumentale oude bomen, zwaar dood hout en kleine openingen in het kronendak een grote betekenis voor de biodiversiteit (Joseph Zwaenepoel bosreservaat, Zoniënwoud) (Foto Luc De Keersmaeker)
Eén soortengroep waar vaak wordt naar verwezen als het over de soortenarme beukenbossen gaat, zijn de vaatplanten (kruiden, varens en paardestaarten). Beukenbossen, zeker op de zuurdere bodems, hebben door de hoge schaduwdruk en het traag afbrekende strooisel vaak weinig ondergroei. Als het aandeel van beuk in een bos toeneemt, neemt het aantal soorten vaatplanten meestal af (Mölder et al., 2008; Mölder et al., 2014). Het gaat dan vooral over lichtminnende soorten, bv. framboos of bleke zegge, die beschikken over een zaadbank of over goede verspreidingsmogelijkheden en die indicatoren zijn van verstoring (Schmidt et al., 2009). Als er een gat in het kronendak ontstaat, komen ze meestal ook snel weer terug tevoorschijn (Degen et al., 2010). Grote openingen, die van nature slechts sporadisch voorkomen in beukenbos, zijn vaak soortenarmer dan kleine openingen omdat ze gedomineerd worden door forse competitieve soorten, zoals bramen (Degen et al. 2010) of adelaarsvaren (Baeté & De Keersmaeker, 2002). Als op voldoende grote schaal gekeken wordt, blijkt het aantal soorten vaatplanten in structuurrijke, onbeheerde beukenbossen minstens even hoog te zijn als in beheerde beukenbossen waar de beheerder regelmatig gaten in kapt (Kaufmann et al., 2018).
Het aantal soorten vaatplanten op een kleine oppervlakte wordt daarom niet als een goede indicator beschouwd voor de natuurwaarde van bossen en de (lokale) achteruitgang van vaatplanten die wijzen op verstoring hoeft dus niet noodzakelijk problematisch te zijn (Boch et al., 2013). Schaduwtolerante oudbosplanten, zoals bosanemoon en wilde hyacint, zijn wel goede kwaliteitsindicatoren en kunnen in gesloten beukenbossen met een hoge schaduwdruk aanzienlijk toenemen (Van Calster et al., 2007). Een tapijt van wilde hyacint, zoals in het Hallerbos, is in belangrijke mate toe te schrijven aan het dichte en gelijkjarige scherm van vitale beuken, dat weinig licht doorlaat (Fig. 6).

Figuur 6: Door de sterke schaduwdruk in gesloten beukenbossen kunnen voorjaarsbloeiers zoals wilde hyacint er tot dominantie komen (Harras, Zoniënwoud; foto Luc De Keersmaeker)
Conclusies en aanbevelingen
De wetenschappelijk literatuur over de positie van beuk in het veranderende klimaat is niet eenduidig en laat nog veel vragen onbeantwoord. Het zou daarom weinig verstandig zijn om hier verregaande beheerconsequenties aan te koppelen en de beuk af te schrijven of beukenbossen versneld om te vormen. Beuk is een schaduwtolerante climaxboomsoort, die minder goed bestand is tegen een combinatie van hoge temperatuur, hoge lichtintensiteit en lage luchtvochtigheid dan lichtbehoevende soorten, zoals bij voorbeeld eiken. Dat maakt de soort vooral in het open landschap en langs bosranden gevoelig voor langdurige droogte en hitte, zoals we die in de voorbije jaren hebben gekend. In gesloten bossen op een goed doorlatende bodem vertoont beuk voorlopig niet meer problemen dan andere soorten en daar verjongt de soort nog steeds succesvol. Als we sterfte vaststellen in bossen, houdt dat vaak verband met de groeiplaats of met beheeringrepen.
Op een ondiepe bodem, die afwisselend nat is en sterk uitdroogt, wortelt beuk ondiep en is de soort gevoelig voor droogtestress en windworp. Mogelijk worden die groeiplaatsen door veranderingen in het neerslagpatroon nog minder geschikt en daarom is het niet aan te bevelen om er nog beuken te planten.
Maar ook door beheeringrepen kunnen beuken het moeilijk krijgen. Wanneer in een bos sterk wordt gedund of een kapvlakte wordt gemaakt, dan krijgen de resterende beuken het heel moeilijk omdat de temperatuur en de instraling sterk toenemen en de luchtvochtigheid afneemt. Het meest extreme en opvallende beeld is dat van beukenbomen met ‘schorsbrand’: wanneer beuken plots in de volle zon komen te staan, dringt de zonnewarmte door tot diep in de stam en tast daar het weefsel aan. Maar ook zonder schorsbrand kunnen beuken hierdoor gezondheidsproblemen krijgen. Als we oude beuken kappen in beukenbossen, omdat we er van uit gaan dat ze ‘gedoemd‘ zijn, veroorzaken we zelf de sterke verstoringen die de overige bomen in de problemen kunnen brengen en dreigt een vicieuze cirkel te ontstaan.
Het beheer van onze beukenbossen sluit dus het best zo goed mogelijk aan bij de natuurlijke, kleinschalige dynamiek van deze bossen, die een garantie is voor het behoud van een koel bosklimaat. In natuurlijke beukenbossen staan jonge en oude bomen door elkaar gemengd, zelfs in elkaars schaduw. Jonge beuken in de schaduw groeien traag en wachten tot een oude beuk het loodje legt. De gaten in het kronendak, die door sterfte van een oude beuk ontstaan, zijn van nature meestal klein en worden snel terug opgevuld door jonge beuken die dankzij het extra licht sneller groeien. Een beukenbos is dus op zichzelf een robuust systeem dat zijn eigen microklimaat en zijn rijke typische biodiversiteit in stand houdt. Niets doen is daarbij een optie: beukenbossen zijn immers op vele groeiplaatsen van nature de climaxvegetatie. In beheerde beukenbossen zijn voorzichtige dunningen en kleine openingen in het kronendak, die het microklimaat niet verstoren, aangewezen. Daar is voldoende aandacht voor oude bomen en zwaar dood hout erg belangrijk: ze zijn immers essentiëel voor het behoud van de gespecialiseerde, vaak weinig opvallende biodiversiteit.
Ook bij herbebossingen op goed doorlatende zand- of leembodem kunnen we nog steeds gebruik maken van beuk. Beuk kan bij voorbeeld een geschikte keuze zijn om op een zure bodem invasieve exoten, zoals Amerikaanse eik, onder controle te krijgen. De hoge schaduwdruk van beuk kan immers de verjonging van exoten terugdringen. Ook in functie van dit beheer is het aangewezen om kleinschalig te werken en een groot bestand van bv. Amerikaanse eik heel geleidelijk aan ‘op te rollen’. Zo blijft het vereiste microklimaat voor de aansluitende en verjongende beuken behouden en realiseren we op termijn een ongelijkjarig, structuurrijk bos.
Bij bosuitbreiding denken we misschien niet in de eerste plaats aan beuk. Vaak planten we nieuw bos op bemeste en bekalkte bodem, die geschikt is voor veeleisende soorten zoals linde en waarvoor vele van onze bossen te zuur geworden zijn. Toch hoeven we bij de aanleg van nieuwe bossen beuken ook niet helemaal uit te sluiten: in kleine groepjes bieden ze zeker een meerwaarde. In dat geval verdient het de aanbeveling om gebruik te maken van pioniers, in een ruim plantverband, waartussen climaxsoorten kunnen worden geplant. Deze pioniers zorgen snel voor een lichte schaduw en een microklimaat dat climaxsoorten zoals beuk beschermt (De Keersmaeker, 2019).
Gelieve als volgt citeren:
Luc De Keersmaeker en Kris Vandekerkhove (2020) Schrijven we de beuk te snel af? Bosrevue 88a, 1-16.
ISSN 2565-6953 – Bosrevue 88a
-
Referenties
Aertsen W., Janssen E., Kint V., Bontemps J.D., Van Orshoven J. & Muys B. (2014) Long-term growth changes of common beech (Fagus sylvatica L.) are less pronounced on highly productive sites. Forest Ecology and Management 312: 252–259.
Assmann T., Drees C., Schroder E. & Ssymank A. (2007) Mythos Artenarmut -Biodiversitat von Buchenwaldern. Natur und Landschaft 82: 401-406.
Baeté H. & De Keersmaeker L. (2002) Kruidlaagonderzoek in de kernvlakte van het bosreservaat Kersselaerspleyn. INBO, Bosreservatennieuws 2: 6.
Boch S., Prati D., Müller J., Socher S., Baumbach H., Buscot F., Gockel S., Hemp A., Hessenmöller D., Kalko E., Linsenmair K. E., Pfeiffer S., Pommer U., Schöning I., Schulze E.-D., Seilwinder C., Weisser W., Wells K. & Fischer M. (2013) High plant species richness indicates management-related disturbances rather than the conservation status of forests. Basic and Applied Ecology 14: 496-505.
Bodin J., Badeau V., Bruno E., Cluzeau C., Moisselin J.‐M., Walther G.‐R. & Dupouey J.‐L. (2013) Shifts of forest species along an elevational gradient in Southeast France: climate change or stand maturation? Journal of Vegetation Science 24: 269-283.
Boussemaere P. & Vicca S. (2020) IT’S THE END OF THE WORLD AS WE KNOW IT. Blog Global Change Ecology, Universiteit Antwerpen. https://globalchangeecology.blog/2020/05/25/its-the-end-of-the-world-as-we-know-it/
Brunet J. & Isacsson G. (2009) Restoration of beech forest for saproxylic beetles—effects of habitat fragmentation and substrate density on species diversity and distribution. Biodiversity and Conservation 18: 2387–2404.
Christensen M., Heilmann‐Clausen J., Walleyn R. & Adamčik S. (2005) Wood‐inhabiting fungi as indicators of nature value in European beech forests. In : Marchetti M. (ed.) Monitoring and indicators of forest biodiversity in Europe—from ideas to operationality. EFI Proceedings 51: 229‐237.
Daise J., Claessens H. & Rondeux J. (2009). Etude de l’adéquation des essences aux stations forestières de la forêt de Soignes (Zone bruxelloise) dans le contexte du changement climatique. Rapport final. ULg – GxABT.
Deforce K., Vanmontfort B. & Vandekerkhove K. (2018) Early and High Medieval (c. 650 AD–1250 AD) Charcoal Production and Its Impact on Woodland Composition in the Northwest-European Lowland: A Study of Charcoal Pit Kilns from Sterrebeek (Central Belgium). Environmental Archaeology, DOI: 10.1080/14614103.2018.1538087
Degen T., Devillez F. & Jacquemart A.-L. (2010) Gaps promote plant diversity in beech forests Luzulo-Fagetum, North Vosges, France. Silvae Genetica 59: 175–182.
De Keersmaeker L. (2019) Advies over de ontwikkeling van ecologisch waardevolle bossen op landbouwgronden. Instituut voor Natuur- en Bosonderzoek, INBO.A.3803.
Dupin A., Girardclos O., Fruchart C., Laplaige C., Nuninger L., Dufraisse A. & Gauthier E. (2017) Anthracology of charcoal kilns in the forest of Chailluz (France) as a tool to understand Franche-Comte forestry from the mid-15th to the early 20th century AD. Quaternary International 458: 200-213.
Dyderski M.K., Paz S., Frelich L.E. & Jagodzinski A.M. (2018) How much does climate change threaten European forest tree species distributions? Global Change Biology 24: 1150-1163. https://onlinelibrary.wiley.com/doi/abs/10.1111/gcb.13925
Everts T., Boeraeve M., Vandekerkhove K., De Keersmaeker, L., Van de Kerckhove P. & Jacquemyn H. (2020) Verjonging van beuk in het Zoniënwoud: terug van weggeweest door ectomycorrhizale schimmels? Natuur.Focus 19(3): 124-131.
Falk W. & Hempelmann, N. (2013) Species favourability shift in europe due to climate change: a case study for Fagus sylvatica L. and Picea abies (L.) Karst. Based on an ensemble of climate models. J. Climatol. 1–18.
Fritz Ö. (2009)Vertical distribution of epiphytic bryophytes and lichens emphasizes the importance of old beeches in conservation. Biodiversity and Conservation 18: 289–304.
Fritz Ö., Niklasson M. & Churski M. (2009) Tree age is a key factor for the conservation of epiphytic lichens and bryophytes in beech forests. Applied Vegetation Science 12: 93–106.
Fritz Ö. & Heilmann-Clausen J. (2010) Rot holes create key microhabitats for epiphytic lichens and bryophytes on beech (Fagus sylvatica). Biological Conservation 143: 1008–1016.
Gessler A., Bottero A., Marshall J. & Arend M. (2020) The way back: recovery of trees from drought and its implication for acclimation. New Phytologist., doi.org/10.1111/nph.16703.
Grebenc T., Christensen M., Vilhar U., Čaater M., Martin M.P., Simoniči P. & Kraigher H. (2009) Response of ectomycorrhizal community structure to gap opening in natural and managed temperate beech-dominated forests. Canadian Journal of Forest Research 39:1375–1386.
Hafner B.D., Hesse B.D. & Grams T.E.E.(2020) Friendly neighbours: Hydraulic redistribution accounts for one quarter of water used by neighbouring drought stressed tree saplings. Plant Cell Environ. 2020: 1– 14.
Härdtle W., Niemeyer T., Assmann T., Baiboks S., Fichtner A., Friedrich U., Lang A.C., Neuwirth B., Pfister L., Ries C., Schuldt A., Simon N. & von Oheimb G. (2013) Long-term trends in tree-ring width and isotope signatures (13C, 15N) of Fagus sylvatica L. on soils with contrasting water supply. Ecosystems 16: 1413–1428.
Heilmann-Clausen J. & Christensen M. (2003) Fungal diversity on decaying beech logs. Implications for sustainable forestry. Biodiversity and Conservation 12: 953-973.
Kaufmann S., Hauck M. & Leuschner C. (2018) Effects of natural forest dynamics on vascular plant, bryophyte, and lichen diversity in primeval Fagus sylvatica forests and comparison with production forests. Journal of Ecology 106: 2421– 2434.
Klein T., Siegwolf R. T. & Körner C. (2016). Belowground carbon trade among tall trees in a temperate forest. Science, 352(6283): 342–344.
Kutsch W.L., Wirth C., Kattge J., Nöllert S., Herbst M. & Kappen L. (2009) Ecophysiological characteristics of mature trees and stands – consequences for old-growth forest productivity. In Wirth C. et al. (eds.) Old-Growth Forests. Ecol. Studies 207. Springer, Berlin, Heidelberg. pp. 57-79.
Lachat T., Wermelinger B., Gossner M.M., Bussler H., Isacsson G. & Müller J. (2012) Saproxylic beetles as indicator species for dead‐wood amount and temperature in European beech forests. Ecological Indicators 23:323–331.
Langohr R. (2009) Hoofdstuk 20: Het Zoniënwoud, uniek voor aardwetenschappen en archeologie. In: De vrienden van het Zoniënwoud (red.): Het Zoniënwoud, een nieuwe visie op een patrimonium met toekomst. Uitgeverijen Mardaga & Groeninghe.
Latte N., Lebourgeois F. & Claessens H. (2015). Increased tree-growth synchronization of beech (Fagus sylvatica L.) in response to climate change in northwestern Europe. Dendrochronologia 33: 69-77.
Leberecht M., Dannenmann M., Gschwendtner S., Bilela S., Meier R., Simon J., Rennenberg H., Schloter M. & Polle A. (2015) Ectomycorrhizal communities on the roots of two beech (Fagus sylvatica) populations from contrasting climates differ in nitrogen acquisition in a common environment. Appl. Env. Microbiol. 81: 5957-5967.
Leuschner C. (2020) Drought response of European beech (Fagus sylvatica L.) – a review,Perspectives in Plant Ecology, Evolution and Systematics, doi:https://doi.org/10.1016/j.ppees.2020.125576
Leuschner, C. & Ellenberg, H. 2017. Ecology of Central European Forests. Springer Nature, Cham.
Maiorano L., Cheddadi R., Zimmermann N.E., Pellissier L., Petitpierre B., Pottier J., Laborde H., Hurdu B.I., Pearman P.B., Psomas A., Singarayer J.S., Broennimann O., Vittoz P., Dubuis A., Edwards M.E., Binney H.A. & Guisan A. (2013) Building the niche through time: using 13,000 years of data to predict the effects of climate change on three tree species in Europe. Global Ecology and Biogeography 22: 302-317.
Máliš F., Bobek P., Hédl R., Chudomelová M., Petřík P., Ujházy K., Ujházyová M. & Kopecký M. (2020) Historical charcoal burning and coppicing suppressed beech and increased forest vegetation heterogeneity. Journal of Vegetation Science, https://doi.org/10.1111/jvs.12923.
Meier E.S., Lischke H., Schmatz D.R. & Zimmermann N.E. (2012) Climate, competition and connectivity affect future migration and ranges of European trees. Global Ecology and Biogeography 21 : 164–178.
Meier I.C., Knutzen F., Eder L.M. et al. Müller-Haubold H., Goebel M.-O., Bachmann J., Hertel D. & Leuschner C. (2018) The Deep Root System of Fagus sylvatica on Sandy Soil: Structure and Variation Across a Precipitation Gradient. Ecosystems 21: 280–296.
Meyer P. & Schmidt M. (2008) Aspekte der Biodiversität von Buchenwäldern – Konsequenzen für eine naturnahe Bewirtschaftung. In: NW-FVA (Hrsg.): Ergebnisse angewandter Forschung zur Buche. Beiträge aus der NW-FVA, Band 3, 159-192 Methodische Empfehlungen. IHW-Verlag, Eching.
MIRA, 2015. Klimaatrapport 2015: over waargenomen en toekomstige klimaatveranderingen. Vlaamse Milieumaatschappij, 2015
Mölder A., Bernhardt-Römermann M. & Schmidt W. (2008) Herb-layer diversity in deciduous forests: raised by tree richness or beaten by beech? Forest Ecology and Management 256:272–281.
Mölder A., Streit M. & Schmidt W. (2014) When beech strikes back: how strict nature conservation reduces herb‐layer diversity and productivity in central European deciduous forests. Forest Ecology and Management 319: 51– 61.
Moning C. & Müller J. (2009). Critical forest age thresholds for diversity of lichens, molluscs and birds in temperate beech (Fagus sylvatica L.) plant communities. Ecological Indicators 9: 922– 932.
Müller J., Brunet J., Brin A., Bouget C., Brustel H., Bussler H., Förster B., Gunnar I., Köhler F., Thibault L. & Gossner M.M. (2012) Implications from large-scale spatial diversity patterns of saproxylic beetles for the conservation of European Beech forests. Insect Conservation and Diversity doi: 10.1111/j.1752-4598.2012.00200.x.
Nabuurs G.J., & Hommel P.W.F.M. (2007) Klimaatverandering en het Nederlandse bos: geen doemscenario’s graag. Vakblad Natuur Bos Landschap 4(8): 8-13.
Nickel U.T., Weikl F., Kerner R., Schäfer C., Kallenbach C., Munch J.C. & Pritsch, K. (2018) Quantitative losses vs. qualitative stability of ectomycorrhizal community response to 3 years of experimental summer drought in a beech-spruche forest. Global Change Biology 24: 560-576.
Ódor P., Heilmann-Clausen J., Christensen M., Aude E., van Dort K.W., Piltaver A., Siller I., Veerkamp M.T., Walleyn R. & Standovar T. (2006) Diversity of dead wood inhabiting fungi and bryophytes in semi-natural beech forests in Europe. Biological Conservation 131: 58-71.
Peltzer D. (2001) Anpassung antioxidativer Systeme an Licht und Temperatur: Holzige und krautige Pflanzen im Vergleich. PhD thesis, University of Göttingen, Germany.
Pena R. & Polle A. (2014) Attributing functions to ectomcyorrhizal fungal identities in assemblages for nitrogen acquisition under stress The ISME Journal 8: 321–330.
Rog I., Rosenstock N.P., Körner C. & Klein T. (2020) Share the wealth: Trees with greater ectomycorrhizal species overlap share more carbon. Molecular Ecology 29: 2321– 2333. https://doi.org/10.1111/mec.15351
Schmidt I., Leuschner C., Mölder A. & Schmidt W. (2009) Structure and composition of the seed bank in monospecific and tree species-rich temperate broad-leaved forests. Forest Ecology and Management 257: 695-702.
Schreel J.D.M., von der Crone J.S., Kangur O. & Steppe K. (2019) Influence of drought on foliar water uptake capacity of temperate tree species. Forests 10: 562.
Shi L., Guttenberger M., Kottke I. & Hampp R. (2002) The effect of drought on mycorrhizas of beech (Fagus sylvatica L.): changes in community structure, and the content of carbohydrates and nitrogen storage bodies of the fungi. Mycorrhiza 12: 303–311.
Sioen G., Verschelde P. & Roskams P. (2020). Bosvitaliteitsinventaris 2019. Resultaten uit het bosvitaliteitsmeetnet. Rapporten van het Instituut voor Natuur- en Bosonderzoek 2020 (20). Instituut voor Natuur- en Bosonderzoek, Brussel.
Speight M. (1989) Saproxylic invertebrates and their conservation. Nature and Environment Series nr. 42. Council of Europe. Strasbourg.
Thurm E.A., Hernandez L., Baltensweiler A., Ayan S., Rasztovits E., Bielak K., Zlatanov T.M., Hladnik D., Balic B., Freudenschuss A., Buchsenmeister R., Falk W. (2018) Alternative tree species under climate warming in managed European forests. Forest Ecology and Management 430: 485-497. https://doi.org/10.1016/j.foreco.2018.08.028
Tolksdorf J.F., Elburg R., Schröder F., Knapp H., Herbig C., Westphal T., et al Schneider B., Fülling A. & Hemker C. ( 2015) Forest exploitation for charcoal production and timber since the 12th century in an intact medieval mining site in the Niederpöbel Valley (Erzgebirge, Eastern Germany). Journal of Archaeological Science: Reports 4: 487–500.
Van Calster H., Baeten L., De Schrijver A., De Keersmaeker L., Rogister J.E., Verheyen K., Hermy M. (2007) Management driven changes (1967 – 2005) in soil acidity and the understory plant community following conversion of a coppice-withstandards forest. Forest Ecology and Management 241: 258-271.
Vandekerkhove K., Baeté H., Van Der Aa B., De Keersmaeker L., Thomaes A., Leyman A. & Verheyen K. (2016) 500 years of coppice-with-standards management in Meerdaal Forest (Central Belgium). iForest 9: 509-517.
Vandekerkhove K., Vanhellemont M., Vrska T., Meyer P., Tabaku V., Thomaes A., Leyman A., De Keersmaeker L. & Verheyen K. (2018a) Very large trees in a lowland old-growth beech (Fagus sylvatica L.) forest: Density, size, growth and spatial patterns in comparison to reference sites in Europe., Forest Ecology and Management 417: 1-17.
Vandekerkhove, K, Deforce, K & Bastiaens, J (2018b) Historic-ecological position of beech in the area of the Sonian Forest and an overview of beech-forest- related biodiversity present in the forest. Rapporten van het Instituut voor Natuur- en Bosonderzoek nr. 29, Instituut voor Natuur- en Bosonderzoek.
Von Arx G., Graf Pannatier E., Thimonier A. & Rebetez M. (2013) Microclimate in forests with varying leaf area index and soil moisture: potential implications for seedling establishment in a changing climate. Journal of Ecology 101: 1201–1213.
Vayreda J., Martinez‐Vilalta J., Gracia M., Canadell J.G. & Retana J. (2016) Anthropogenic‐driven rapid shifts in tree distribution lead to increased dominance of broadleaf species. Global Change Biology 22: 3984-3995.
Zellweger F., De Frenne P., Lenoir J., Vangansbeke P., Verheyen K., Bernhardt‐Römermann M., … & Coomes D. (2020). Forest microclimate dynamics drive plant responses to warming. Science, 368: 772– 775.
Zohner C. M., Mo L., Renner S. S., Svenning J.‐C., Vitasse Y., Benito B. M., … Crowther, T. W. (2020). Late-spring frost risk between 1959 and 2017 decreased in North America but increased in Europe and Asia. Proceedings of the National Academy of Sciences Jun 2020, 117 (22) 12192-12200; DOI: 10.1073/pnas.1920816117


