


Bosrevue
De jonge en dichte bosfase vormen samen een vroege periode van bosontwikkeling waarin een nieuwe generatie bomen zich succesvol heeft kunnen vestigen maar de opstand nog niet of nog maar net in kroonsluiting is. Deze vroege ontwikkelingsfasen zijn bijzonder dynamisch door het ontstaan van allerhande interacties tussen bomen en begeleidende vegetatie en naderhand tussen bomen onderling. Sommige van deze interacties zijn voordelig, zoals de vermindering van de gevoeligheid voor vraat en ongunstige weersomstandigheden. Andere interacties zijn dan weer nadelig, vooral wanneer bekeken vanuit het perspectief van een individuele boom. Groei en ontwikkeling leidt namelijk in toenemende mate tot ondergrondse competitie voor water en voedingsstoffen en bovengrondse competitie voor licht en ruimte. Bomen reageren op competitie door o.a. hun groeipatroon voortdurend aan te passen aan de omgeving. Denk voorbeeld aan randbomen die asymmetrisch groeien in de richting met het meeste licht.
Flexibele groei en ontwikkeling van bosbomen
De vormontwikkeling van bosbomen heeft een belangrijke invloed op de stamkwaliteit (Kint et al. 2010). Zo hebben vitale bomen met een rechte en lange takvrije stam veelal het grootste kwaliteitspotentieel voor latere toepassingen met hoge toegevoegde waarde, zoals zaag- of fineerhoutproductie. De boomvormontwikkeling hangt in eerste instantie af van soortkenmerken, genetische eigenschappen en groeiplaatseigenschappen. Maar ze wordt verder ook beïnvloed door lichtcompetitie. Deze ecologische kennis vormt al van oudsher de basis van vele bosbeheersystemen. Zo wordt er vaak eerst aandacht besteed aan de ontwikkeling van rechte stammen, dunne zijtakken en vitale kruinen. Het behouden van een hoge competitiedruk zal er nadien voor zorgen dat dunne zijtakken afsterven en afvallen. Zodra natuurlijke takreiniging zich voldoende hoog heeft voltrokken (het zogenaamde omslagpunt), worden de bomen vrijgesteld om een maximale diameteraanwas te bekomen. Hoewel boomvormontwikkeling een basisbegrip is in het bosbeheer, blijven er grote kennishiaten.
Kennishiaten over gemengde jonge bosopstanden
Ten eerste is veel praktijkervaring en wetenschappelijke kennis gebaseerd op observaties in monoculturen. De keuze voor monoculturen staat vandaag de dag onder discussie, ondermeer omwille van hun beperkte stabiliteit, met name in het licht van klimaatverandering, en lagere multifunctionaliteit (Nadrowski, Wirth & Scherer-Lorenzen 2010). De mogelijkheid om ook hoogkwalitatieve stammen te produceren in soortenrijke bossen kan de aanplant ervan verder motiveren. Maar verloopt architecturale ontwikkeling, als basis voor de productie van hoogkwalitatieve stammen, hetzelfde als in monocultuur? Wellicht niet. De invloed van competitie op boomvormontwikkeling hangt af van de competitiekracht en competitietolerantie van aanwezige soorten in de menging, en is daarom niet volledig voorspelbaar vanuit ervaringen met monoculturen (Dieler & Pretzsch 2013).
De tweede kennishiaat gaat over het moment waarbij boomarchitectuur een relevante maatstaf is voor de uiteindelijke boomkwaliteit. In literatuur wordt vooral de stakenfase beschreven, een moment waarop een 6-8 meter takvrije onderstam quasi volledig ontwikkeld is (Kantola, Mäkinen & Mäkelä 2007; Hein 2008). Ook in de praktijk worden toekomstbomen op dat moment geselecteerd. Echter, groei en ontwikkeling tijdens de jonge en dichte bosfasen hebben mogelijk een bepalende, en deels onomkeerbare, invloed gehad op de stamvorm. Een minder voorkomend alternatief is dan ook om toekomstboom-opties te selecteren in een vroeger stadium van bosontwikkeling. Een deel van deze opties zullen nadien doorgroeien tot volwaardige toekomstbomen. Het voordeel van de strategie is een maximaal kwaliteitspotentieel door boomvorm vanaf het begin op te volgen, en eventueel ook te sturen. Het nadeel is dat een groot aantal bomen geëvalueerd moet worden, wat dan weer een extra tijdsinvestering is.
Doelstelling van het onderzoek
Om boomvormontwikkeling en kwaliteitspotentieel in jonge, gemengde bosopstanden beter te begrijpen, zochten we antwoorden op vier onderzoeksvragen:
- Vraag 1: Heeft competitie een invloed op de vormontwikkeling van jonge bosbomen?
- Vraag 2: Verloopt vormontwikkeling gelijkaardig in gemengde en ongemengde opstanden?
- Vraag 3: Welke omgevingskenmerken en boomvormkenmerken zijn bepalend voor de kwaliteitsbeoordeling van jonge bosbomen?
- Vraag 4: Wat zijn de percepties van bosbeheerders over zo’n vroege kwaliteitsbeoordeling?
Dataverzameling in FORBIO
Het boomsoortendiversiteitsexperiment FORBIO-Zedelgem (zie bosrevue 54) is geschikt om deze onderzoeksvragen te beantwoorden. In 2010 werd zeven hectare landbouwgrond beplant met ongeveer 33 000 bomen van vijf soorten (berk, beuk, grove den, winterlinde en zomereik) met een plantafstand van anderhalve meter. Deze bomen groeien in monoculturen en in verschillende twee-, drie- en viersoortenmengingen. Het experiment ondergaat op dit moment een overgangsfase van jong naar dicht bos met een gemiddelde kroonsluiting van 63 % (Fig.1). De gemiddelde boomhoogte tijdens de meetcampagne (in de zomer van 2015) bedroeg 210 cm, behalve voor berk die al een gemiddelde hoogte van 480 cm had bereikt.
De bovengrondse architectuur van 400 bomen werd in detail opgemeten. Hiervoor werden diameter, lengte, inplantingshoek, richting en hoogte van alle zijtakken op de hoofdstam bepaald. Verder werden afwijkingen zoals scheefgroei, kronkelende stamvorm, apicale dominantie, en kruinvitaliteit genoteerd. Op basis van de soort en de grootte van buurbomen werd de groeiomgeving van iedere boom vastgelegd. Deze informatie volstaat alvast om onderzoeksvragen 1 en 2 te beantwoorden.

Figuur 1: De overgangsfase van jong naar dichte bos in FORBIO-Zedelgem in 2015. Links: het midden van een zone met zomereik en berk; rechts: de rand van een zone met grove den, zomereik en berk.
Vormontwikkeling in functie van competitie en menging (Vragen 1 en 2)
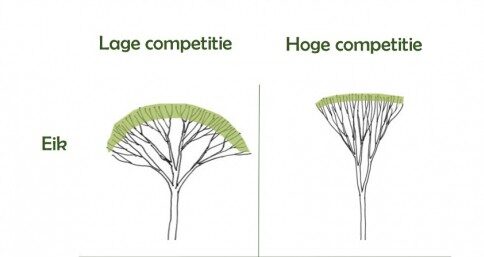
De jonge opstand is nog niet volledig in kroonsluiting maar boomvormontwikkeling wordt al beïnvloed door competitie (Fig. 2). Berk is bijvoorbeeld een snelgroeiende en lichtbehoevende soort, en bij toenemende competitie (beschaduwing) is de investering in hoogtegroei belangrijker dan de investering in diametergroei. Gelijktijdig zullen de onderste takken afsterven omdat fotosynthese niet meer rendabel is. Slechts vijf jaar na aanplant is natuurlijke takreiniging voor berk dus al ingezet. Zomereik is een relatief lichtbehoevende soort, maar in tegenstelling tot berk verloopt de jeugdgroei trager. Bij toenemende competitie zal zomereik meer investeren in hoogtegroei dan in stamdiametergroei en gaan de takken ook scherper omhoog gericht worden. Beide architecturale aanpassingen hebben als voordeel dat er meer licht beschikbaar is voor fotosynthese. Anderzijds zullen scherp ingeplante takken een hogere apicale dominantie opbouwen, sneller dik te worden en dus moeilijker afsterven. Beuk kan zich ten slotte goed handhaven onder schaduw, maar zal onder deze omstandigheden ook meer zijtakken produceren (zie ook Rozenbergar & Diaci 2014). Dat betekent dat beuk de concurrentie voor licht en ruimte met buurbomen zal aangaan omdat de soort bij lage lichtintensiteit een competitief voordeel heeft.
De vormontwikkeling van bosbomen wordt dus vanaf een vroege ontwikkelingsfase beïnvloed door competitie, maar verloopt dit proces gelijkaardig in gemengde en ongemengde opstanden? Onze resultaten tonen aan dat zo’n verschil nog niet meetbaar was. Ook competitie veroorzaakt door lichtbehoevende of schaduwtolerante soorten kan niet van elkaar onderscheiden worden. Er is één uitzondering: berk. Omdat de jeugdgroei van berk ongeveer twee keer zo sterk is als die van de andere soorten, is competitie in een berkenmonocultuur beduidend hoger dan in menging met andere soorten. Zoals net besproken leidt dit tot slankere bomen en minder levende zijtakken. Aangezien processen dus sneller verlopen in berken-monocultuur, zullen ook beheeracties elkaar snel opvolgen.
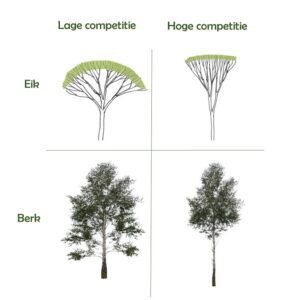
Figuur 2: Illustratie over hoe zomereik en berk zich ontwikkelen onder hoge en lage competitie. Hoge competitie zorgt voor grotere investering in hoogtegroei t.o.v. diametergroei (beide soorten), omhooggerichte takken (zomereik) en natuurlijke takreiniging (berk). Deze figuur is een bewerking uit Pretzsch (2014).
Kwaliteitsbeoordeling van jonge bosbomen (Vraag 3)
Om antwoord te geven op onderzoeksvraag 3 werd het kwaliteitspotentieel van bomen gescoord. Dat gebeurde in juni 2016 tijdens de Pro Silva excursie FORBIO: experimenteren met soortenmengingen. Beheerders van private en openbare bossen werden uitgenodigd om het proefveld in Zedelgem te bezoeken en om deel te nemen aan een wetenschappelijke oefening. In kleine groepjes bewandelden 21 deelnemers verschillende trajecten doorheen het experiment en passeerden daarbij de bomen met gekende architectuur. Er werd hen gevraagd om de kwaliteit van deze bomen in te schatten op basis van vier categorieën (geen, weinig, voldoende en hoog kwaliteitspotentieel). In totaal werden ongeveer 1087 kwaliteitsscores van 211 bomen verzameld.
De resultaten (Tabel 1) tonen aan dat boomkwaliteit negatief beïnvloed wordt door het kronkelend verloop van de hoofdstam, scheefgroei en het verlies van apicale dominantie (i.e. het fenomeen waarbij één of meerdere zijtakken zich evenwaardig gedragen als de hoofspil; vaak met een gesplitste stamvorm of dikke zijtakken als gevolg). Een kronkelend verloop van de hoofdstam verdwijnt bij sommige soorten gemakkelijk met het verdikken van de stam (bv. beuk). Scheefgroei en verlies van apicale dominantie hebben vaker een blijvende impact, en bij oudere bomen zijn deze vormproblemen ook moeilijk te corrigeren via beheer. Verlies van apicale dominantie was vooral bij grove den en winterlinde een probleem en kan te wijten zijn aan bebossing op voormalig landbouwgrond of aan de gebruikte herkomst. Opvallend is dat andere architecturale kenmerken, zoals takdiameter, inplantingshoek en het aantal zijtakken, niet belangrijk waren bij de beoordeling van toekomstige boomkwaliteit.
De gemiddelde kwaliteitsscore van soorten in gemengde en ongemengde opstanden verschilt niet van elkaar. Wanneer geëvalueerd tijdens de jonge ontwikkelingsfase blijven dus alle kansen open om gelijkwaardige houtproducten te produceren in gemengde en ongemengde bestanden. Boomkwaliteit werd wel aanzienlijk lager ingeschat in omgevingen met een hoge competitiedruk (Fig. 3). Er zijn twee mogelijke verklaringen. Ofwel wordt de boomvorm effectief minder gunstig ontwikkeld bij intense beschaduwing. Echter, vormkenmerken die de kwaliteitsbeoordeling bepalen (kronkelend verloop van de hoofdstam, scheefgroei en verlies van apicale dominantie) bleken niet beïnvloed te worden door competitie. Het is daarom waarschijnlijker dat een boom door de deelnemers in combinatie met zijn directe groeiomgeving werd beoordeeld. Dit wil zeggen, bomen die opgroeien in een omgeving met een relatief hoge competitiedruk hebben een groter risico om onderstandig te worden ten opzichte van buurbomen. Deze onderstandige bomen zijn veelal geen goede kandidaat-toekomstbomen.
Tabel 1: De boomkwaliteitsscores van bosbeheerders kunnen verklaard worden aan de hand van boomvormkenmerken in een cumulatief gemengd regressiemodel. Variabelen in het vet hebben een belangrijke invloed op de kwaliteitsscore (significantieniveau p < 0.01).

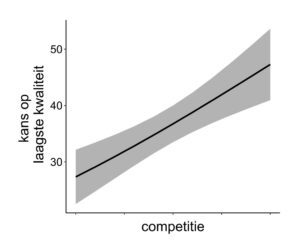
Figuur 3: Kans op laagste kwaliteitsbeoordeling van jonge bosbomen neemt toe in omgevingen met een hoge competitiedruk veroorzaakt door buurbomen.
Percepties over de kwaliteitsbeoordeling (vraag 4)
De variatie in kwaliteitsbeoordeling tussen de deelnemers was groot, maar die variatie kon niet verklaard worden aan de hand van profielkenmerken (zoals leeftijd, geslacht, ervaring in bosbeheer, ervaring met kwaliteitsbeoordeling van toekomstbomen en het onderscheid tussen privé en openbaar beheer). De kwaliteit werd ook algemeen laag inschat (Fig.4). Tijdens de groepsdiscussie bleek dat men de opstanden te jong vond voor zo’n kwaliteitsinschatting. Men verwacht dat de huidige boomkenmerken sterk zullen veranderen en daarmee slechts in beperkte mate representatief zijn voor toekomstige kwaliteit. De economische rendabiliteit voor het selecteren van toekomstboomopties werd dan ook in vraag gesteld. Men verwacht dat er zonder vroegtijdige beheerinterventies nog voldoende kwaliteitsvolle bomen zullen overblijven. Dat zou leiden tot een uitgespaarde kost voor het vroegtijdig aanduiden en eventueel behandelen van een groot aantal toekomstboom-opties. Indien nodig kan de kwaliteit van een beperkt aantal toekomstbomen in een latere bosontwikkelingsfase nog verbeterd worden, bijvoorbeeld door handmatige opsnoei. De instandhouding van soortenmenging op een kleine ruimtelijke schaal was volgens de deelnemers wel een belangrijke reden om bosontwikkeling vanaf een vroege fase op te volgen. De aanplant in het FORBIO-experiment gebeurde immers in blokken met 9 bomen van eenzelfde soort. Grote verschillen in jeugdgroei kunnen er dan voor zorgen dat de minst dominante soorten wegconcurreert worden en dat de gewenste soortenmenging niet behouden blijft.
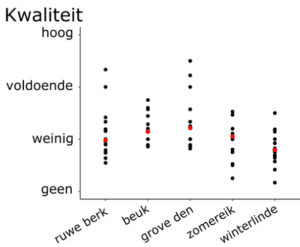
Figuur 4: Kwaliteitsbeoordeling van individuele personen (zwart) en algemeen gemiddelde (rood) voor de 5 boomsoorten in het experiment.
Toekomstvisie
Het FORBIO-experiment is ontworpen om bosbouwkundige vraagstukken in verband met soortenmengingen op een korte en lange termijn te evalueren. De opmeting van boomarchitectuur en de kwaliteitsbeoordeling zullen dan ook opnieuw geëvalueerd worden wanneer de opstanden verder ontwikkeld zijn. Bovendien werd inmiddels een deel van de bomen gesnoeid, en een ander deel niet. Dit zal ons toelaten om op objectieve wijze te evalueren of beheeringrepen tijdens de vroege fase een zinvolle investering zijn.
Besluit
Vraag 1: Heeft competitie een invloed op de vormontwikkeling van jonge bosbomen?
Ja, nog voor volledige kroonsluiting wordt vormontwikkeling van jonge bosbomen beïnvloed door onderlinge lichtcompetitie.
Vraag 2: Verloopt die ontwikkeling gelijkaardig in gemengde en ongemengde opstanden?
Ja, behalve voor berk kan er in de vroege bosontwikkelingsfase nog geen onderscheid gemaakt worden tussen de boomvorm van een soort in gemengde en ongemengde omgeving.
Vraag 3: Welke omgevingskenmerken en boomvormkenmerken zijn bepalend voor de kwaliteitsbeoordeling van jonge bosbomen?
Beoordeling van boomkwaliteit wordt beïnvloed door competitie (maar niet door soortenmenging) en verder door het kronkelend verloop van de hoofdstam, scheefgroei en het verlies van apicale dominantie (maar niet door andere takkenmerken).
Vraag 4: Wat zijn de percepties van bosbeheerders over zo’n kwaliteitsbeoordeling?
Algemeen werd gesteld dat bestanden voor kroonsluiting wellicht te jong zijn om toekomstige kwaliteitskansen te beoordelen. De economische rendabiliteit voor het vroegtijdig selecteren van toekomstboom-opties werd dan ook in vraag gesteld. Monitoring van de ontwikkeling van (al dan niet gesnoeide) toekomstboom-opties zal de zin of onzin van hun vroegtijdige selectie uitwijzen.
Dankwoord
De auteurs willen BOS+ en het ANB beheerteam in Vloethemveld bedanken voor de organisatie van de Pro Silva excursie naar FORBIO-Zedelgem (juni 2016). Ook alle deelnemers worden bijzonder bedankt voor hun waardevolle bijdrage aan dit onderzoek.
Contactpersonen:
Kris Verheyen (kris.verheyen@ugent.be) en Thomas Van de Peer (thomas.vandepeer@kuleuven.be)
Website FORBIO: www.treedivbelgium.ugent.be
ISSN 2565-6953 – Bosrevue 62a
-
Referenties
Wetenschappelijk artikel waarover gerapporteerd:
Van de Peer T, Verheyen K, Kint V, Van Cleemput E, Muys B (2017) Plasticity of tree architecture through interspecific and intraspecific competition in a young experimental plantation. Forest Ecology and Management 385, 1-9
Andere referenties uit de tekst:
Dieler, J. & Pretzsch, H. (2013) Morphological plasticity of European beech (Fagus sylvatica L.) in pure and mixed-species stands. Forest Ecology and Management, 295, 97–108.
Hein, S. (2008) Knot attributes and occlusion of naturally pruned branches of Fagus sylvatica. Forest Ecology and Management, 256, 2046–2057.
Kantola, A., Mäkinen, H. & Mäkelä, A. (2007) Stem form and branchiness of Norway spruce as a sawn timber-Predicted by a process based model. Forest Ecology and Management, 241, 209–222.
Kint, V., Hein, S., Campioli, M. & Muys, B. (2010) Modelling self-pruning and branch attributes for young Quercus robur L. and Fagus sylvatica L. trees. Forest Ecology and Management, 260, 2023–2034.
Nadrowski, K., Wirth, C. & Scherer-Lorenzen, M. (2010) Is forest diversity driving ecosystem function and service? Current Opinion in Environmental Sustainability, 2, 75–79.
Pretzsch, H. (2014) Canopy space filling and tree crown morphology in mixed-species stands compared with monocultures. Forest Ecology and Management, 327, 251–264.
Rozenbergar, D. & Diaci, J. (2014) Architecture of Fagus sylvatica regeneration improves over time in mixed old-growth and managed forests. Forest Ecology and Management, 318, 334–340.


